Abstract
The genomic era has provided unassailable evidence that humans have evolved from common ancestors we share with chimpanzees and (further back in time) with all other primates and with all other mammals. One class of this evidence is the presence of ancient viral genes that were spliced into the genomes of our prehuman ancestors and transmitted to us. Retroviruses are the classical exemplar of this phenomenon, but more recently genes derived from potentially pathogenic bornaviruses have been discovered in our genome. At least two of these genes have been coopted to provide important functions. The advent of humanity, due in part to capabilities generated by random genetic mechanisms, is describable in theological terms as creatio ex vetere—creation of the new from the old (from stardust and antecedent species). This concept is applicable to the biblical depiction of human development, as seen in the commissioning of humanity as the image of God. Genetic changes are usually innocuous but may generate either disease or new capabilities. The cost of evolution reflects the biblical theme that suffering precedes glory, of which the history of Jesus is paradigmatic. Our biological history argues against our tendency to self-glorification—our hubris—but can be seen, from a theological point of view, to be part of the divine plan by which a redeemed and transformed humanity will be raised to share in the very life of God.
This paper is the third in a series that reflects on the activities of semiautonomous agents that operate within genomes and continuously modify them. These agents act as markers that establish the reality of human evolution. They clarify the way scientific concepts should be distinguished from, but may be interpreted by, theological ones. The first paper[1] shows that multispecies comparisons of genomes are analogous to textual criticism of manuscripts. Genomes and manuscripts evolve over time. The accumulation of variants (genetic and textual) delineates histories. Evolution is history, an aspect of our spacetime, created reality. Creation and evolution cannot be mutually exclusive, as is often claimed. God’s continual upholding of history provides a basis for the concept of creatio continua. The second paper[2] describes how new features arise in genomes, and how random process in the context of lawful constraints is the (God-given) means by which evolutionary change proceeds. Molecular process must be distinguished from the agency of God, which is to give being, existence. This is the basis of our understanding that divine action creates unprecedented realities (traditionally, creation from nothing), creatio ex nihilo.
The current paper discusses a group of genes, present in our DNA, that have been derived from pathogenic (disease-causing) viruses. They demonstrate our continuity with nonhuman mammals. The divine Son became incarnate within this biological matrix, and from it God will resurrect believing humanity at the eschaton. Such transformations relate to the theological idea of creation out of the old, creatio ex vetere.
Inherited Segments of Bornavirus DNA
These are heady times for scientific pathologists. Novel pathogenic viruses affecting people are continuing to appear at regular intervals.[3] Typically, the emergent disease-causing viruses preexist in nonhuman species, in which they do not cause overt disease, and acquire the capacity to infect humans. Such nonhuman-to-human infections are said to be zoonotic. In quick succession, humanity has been challenged with outbreaks of Chikungunya and Zika, MERS, SARS, and COVID19, and latterly unprecedented monkeypox outbreaks. Novel strains of influenza lurk in the wings.
In parts of Germany, bornaviruses have been identified as being responsible for rare but devastating neurological disease of humans. These viruses occur naturally in shrews and (more rarely) in squirrels, in which they are nonpathogenic. But if they infect humans, they cause encephalitis (inflammation) that involves numerous regions of the brain. Encephalitis leads to death or severe permanent disability.[4] Research is afoot to discover the geographic range of this viral disease, and to ascertain whether hitherto unexplained cases of encephalitis have a bornavirus aetiology.
Surprisingly, lengths of bornavirus-derived DNA are found in the human genome—yours, mine, everyone’s. We did not acquire these bornavirus genes from infectious viruses, but we inherited them from our parents, and they from previous generations. Genetically transmitted bornavirus genes are said to be endogenous, and they are added to animal genomes by a random mechanism mediated by enzymes provided by genomic parasites called transposable elements (Figure 1).[5]
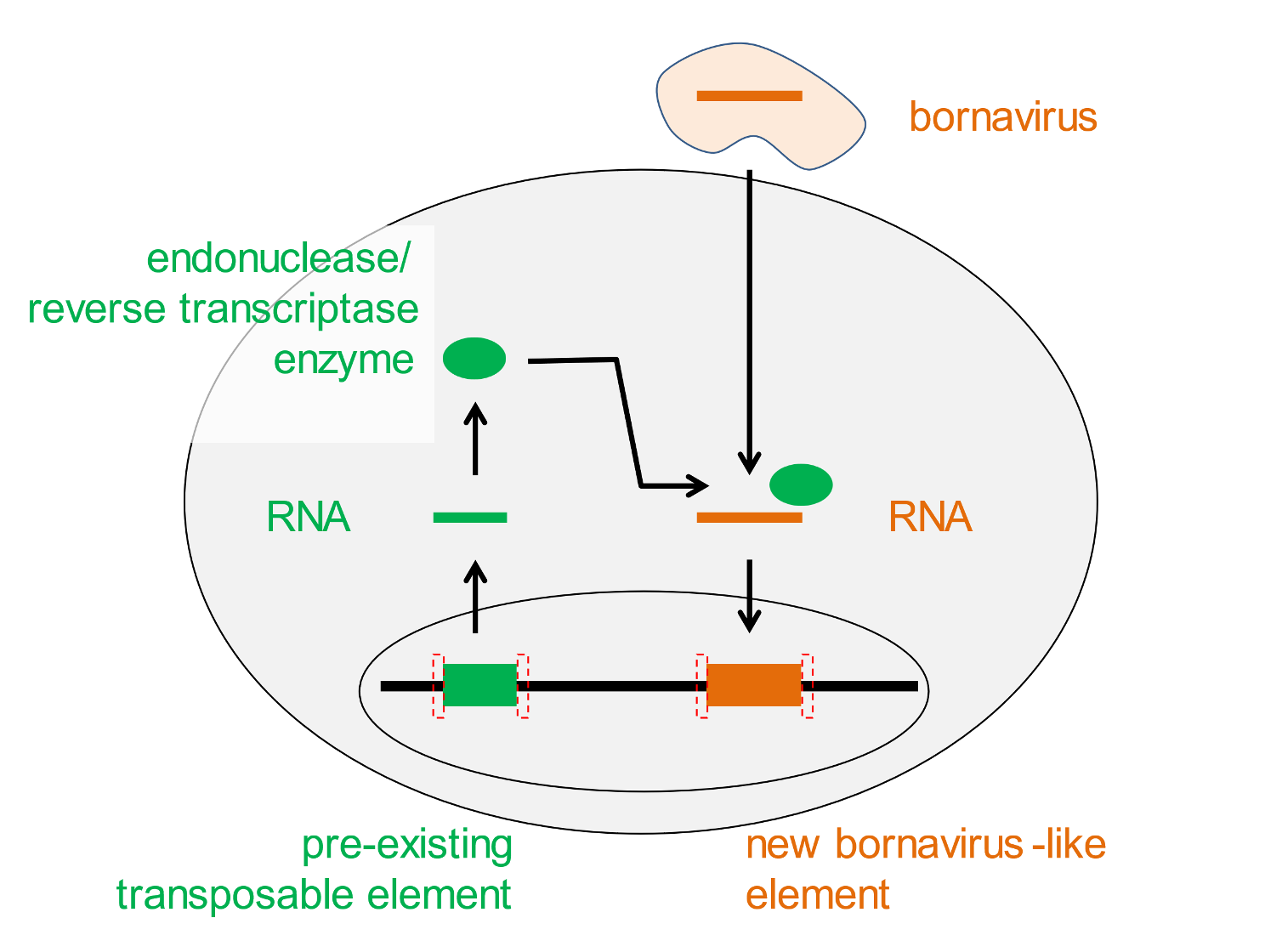
Figure 1. How endogenous bornavirus-like (EBL) elements arise. Bornaviruses lack enzymes to splice their genetic material into chromosomal DNA. Preexisting transposable elements in a cell’s genome generate an RNA molecule that specifies the production of the necessary enzyme (green symbols). These enzymes may fortuitously associate (bent arrow) with one of the RNA molecules that comprises the genome of an infectious bornavirus and insert a DNA copy of the latter into the cell’s genome. A hallmark of this reaction is duplicated target sites that bracket the inserted foreign DNA (red dotted boxes). If this event occurs in a reproductive cell, the randomly acquired endogenised bornavirus gene can be transmitted to future generations.
Given that we all possess the same set of endogenous bornaviral genetic segments, we must all be descended from individuals (more precisely, from their reproductive cells) which hosted bornavirus infections.[6] But when did these ancestors live? Comparative studies of the genomes of many species indicate that the most recent of these acquired segments of bornavirus DNA in the human genome are shared with (other) apes and Old World monkeys (the segments EBLN12 and EBLN22). This demonstrates that humans, other apes, and Old World monkeys are the co-descendants of the individuals (that lived abut forty million years ago) in which each of the respective bornavirus DNA segments arose. We and macaques share the same ancestors. Thirteen instances of bornavirus-derived segments are shared by humans and all simian species; two are shared by all primates; and five by all Boreoeutherian mammals (which include, in addition to primates, orders such as rodents, bats, carnivores, cattle, and whales).[7] Figure 2 depicts the times at which each of twenty-two bornavirus-derived DNA segments were added to the genome we have inherited, and three others to New World monkey genomes.
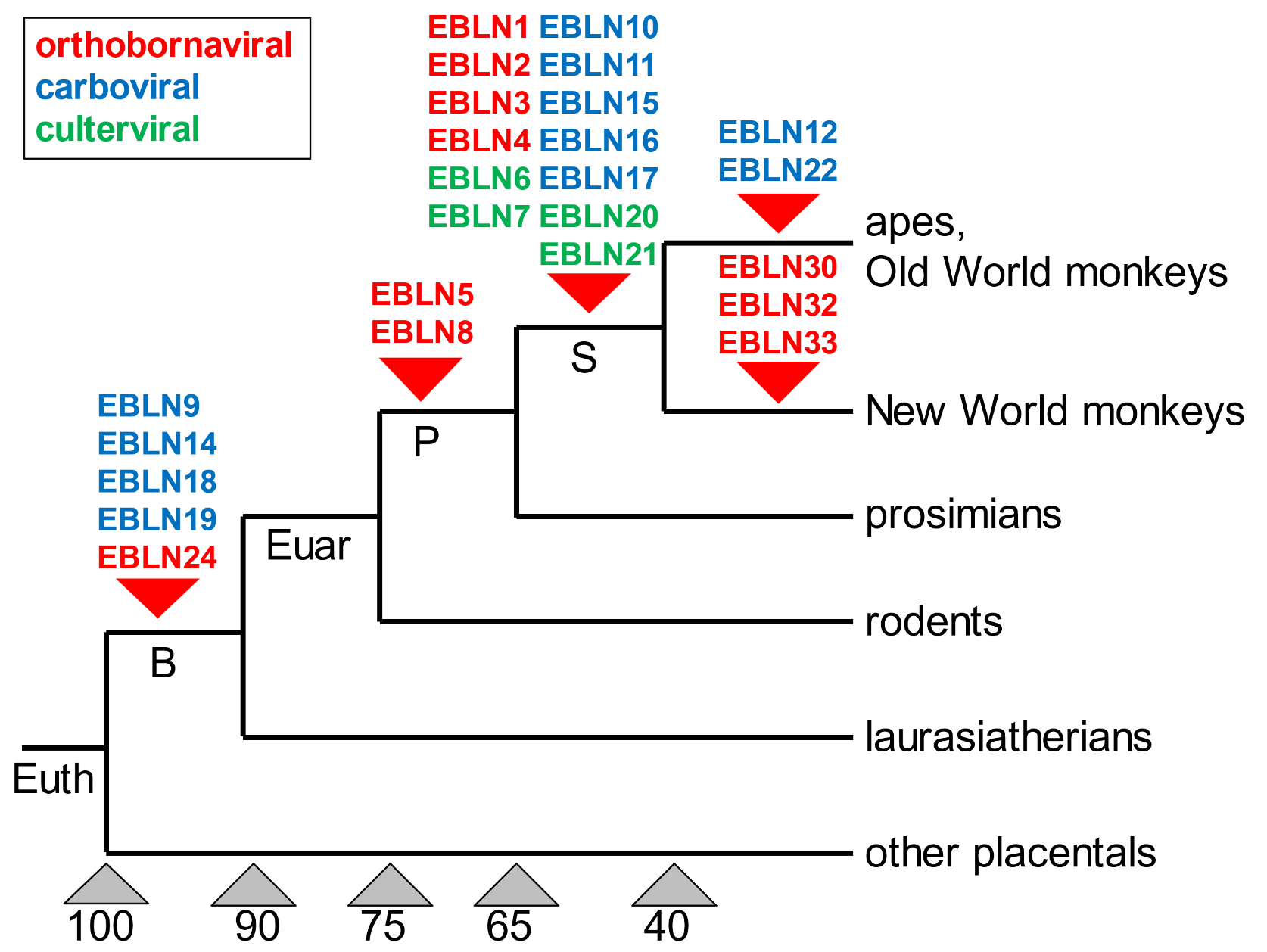
Figure 2. Times at which bornaviral genes were inserted into mammalian genomes. Colours represent three different subtypes of bornavirus. As an example of how to interpret this dendrogram, consider the bornaviral DNA segments known as EBLN5 and EBLN8. These are present in the genomes of all primates but not in the genomes of any other mammals. Each segment was therefore inserted in the genome of a primate ancestor. These data establish that the primates, including humans, are monophyletic, descendants of the same ancestral lineage. Such lineages are given as S, simians; P, primates; Euar, Euarchontoglires (a taxon including primates and rodents); B, Boreoeutherians (the above as well as Laurasiatherian mammals that include moles and hedgehogs, bats, carnivores, hoofed animals, and whales); Euth, Eutherian, all placental mammals (including elephants and armadillos). Numerals at the bottom of the dendrogram indicate million years ago (approximately) when major splits occurred. Adapted from Kawasaki et al. (2021), Figure 3D.
What are theological implications of these endogenous bornavirus genes? When different cells or organisms share uniquely arising mutations (such as insertions of bits of bornavirus-derived DNA), we are permitted only one scientific interpretation of the data: those mutations, which arose in a single event, have been inherited by all the cells or organisms that now possess them. These findings alone are sufficient to demonstrate our evolutionary origins. Humans have inherited mutations that arose millions of years ago, in ancestral species that were very different from our own species. Such genetic findings establish that we are evolved organisms—we have phylogenetic continuity with all other Boreoeutherian mammals. This evidence must bring closure to decades of theologically motivated debate over our evolutionary history.[8]
Our evolutionary history illuminates the nature of our physical embodiment and the theological assertion that we are made of the dust of the earth[9]—the same matter as all other organisms. In scientific terms, human biology is wholly continuous with that of other animals. Our genetic connectedness suggests that many of our physical and temperamental weaknesses have deep origins in biology. Human nature is influenced by an unbroken history connecting us to our primate and other more remote biological antecedents. We need to consider our physicality very seriously, even though as spiritual beings we are qualitatively distinguished from the world of nonhuman animal nature.
In the New Testament, our physical body or flesh (Greek, sarx) may refer to the whole person in all its ambiguity, whether an individual is considered to be morally polluted or purified.[10] Our embodiment as sarx is subject to weakness,[11] to the effects of conflict or fear,[12] and to destructive behaviours, appetites, and instincts.[13] Our inherited sarx, to which reason or moral responsibility has been superadded, underlies and motivates our rebellion against God.[14] Thus, even though sarx refers to our normal embodiment, for Paul the term carries negative connotations, “always pulling down towards decay and death, towards the old creation which is subject to futility.”[15] In John’s gospel, sarx is impotent to transcend itself morally or spiritually.[16] The term flesh denotes either our biological physicality or, by extension, the whole personality of human beings as orientated to self-will and self-gratification.
Strikingly, Jesus the Messiah came to share in this anthropoid primate flesh, this fully embodied humanity,[17] the offering of which was redemptive for all those whose possession of sarx entailed entrapment in sin.[18] Redemption of our embodied being points to the mystery of God’s grace, by which a particular evolved creature, deeply embedded in its biological roots, has been chosen to be liberated from its selfishness and violence, and to be destined to share in the very life and nature of God.[19]
Returning to molecular genetics, we find more surprises that have arisen from bornavirus research. One might expect that foreign segments of DNA, inserted at random into cellular genomes, would be of no use to host organisms. There is no selective pressure to maintain their protein-coding capacity, and with the passage of time they would accumulate mutations and decay into degenerated relics—as most appear to do. However, in at least two cases, particular bornavirus segments in our DNA have retained the capacity to specify the production of proteins, which now serve us, their hosts. The locations of these two genes in the human genome, called endogenous bornavirus-like nucleoprotein 1 and 2 (EBLN1 and EBLN2) have been published, and the precise sites at which they are inserted in the human and other primate genomes are compared below, using the approach described elsewhere.[20]
The genomic location at which the EBLN1 gene resides is depicted in Figure 3. Genetic historiography reveals a wealth of detail as to how this tiny segment of genome has changed over time. Stepwise from the bottom of the diagram we may reconstruct the following sequential developments:
- For species distantly related to us, a small segment (we could call it an “excerpt”) of genetic text is shown, about 42 letters in length. Each letter represents a DNA base, the ultimate unit of hereditary information. This segment of text contains the site into which, in a later ancestor of the simian primates, the bornavirus gene was to be inserted. The uninterrupted site is preserved in prosimians (tarsiers and the aye-aye, galago and lemur group) and in some non-primates (colugos and the horse-rhino-tapir group). It could not be identified in the genomes of many distantly related mammals, in which it has presumably diverged beyond recognition, or has been deleted.
- Using the strategies of textual criticism,[21] we can reconstruct from these variant genetic texts a single original sequence (or, as a geneticist would say, a consensus sequence). This is the sequence that would have occurred in an ancestor of primates and horses.
- During the early history of the primates (but after the tarsier lineage diverged from that leading to simians), four letters (CATT, in red) were deleted. This produced the length of text (the reconstructed target site) into which the foreign bornavirus gene was inserted. This inferred target site has not been identified in any extant species and it has been lost from the genetic record. Only the lineage leading from the bornavirus gene insert has survived.
- All simian primates (humans to capuchin monkeys) possess the inserted gene, although in New World monkeys (marmoset to capuchin monkey), the right hand part of the insert has undergone a deletion. The extreme left and right hand ends of the bornavirus-derived DNA are shown in orange. Hundreds of letters (bases) lie between these termini as indicated by the “…” ellipsis.
The target site (in bold and shaded) is duplicated during the insertion process, and it acts to bracket the foreign viral gene sequence. Target site duplication is a property of an enzyme called an endonuclease/reverse transcriptase, donated fortuitously by parasitic units of DNA that reside in the genome (Figure 1). The mechanism of the insertion, that occurred some forty million years ago, is known. This observation is important because it shows that the insertion event occurred by a familiar molecular process.[22] Natural process is God’s modus operandi in biological history. Two facts demonstrate that every species that possesses the insertion inherited it from the one cell in which it uniquely arose. First, insertion sites are selected at random in the vastness of the genome.[23] Second, the EBLN1 insertion site is at the same location in every species in which it is found. Such a singular addition to the genome could not occur independently in more than one cell. Our evolution from a simian ancestor is confirmed.

Figure 3. Insertion site of EBLN1 gene. DNA sequences are shown for thirteen simian species possessing the insert, and for four prosimian and four non-primate species in which the undisturbed target site could be identified. The ellipsis “…” indicates that DNA sequence extends for millions of bases to the left and right of the segments shown, and (internally) for hundreds of bases comprising the bornaviral DNA insert. In this and later figures, bases shaded and in bold represent the target site and its duplications. All sequences with the undisturbed target site include four bases (typically, CATT), which must have been absent in the cell sustaining the bornaviral DNA insertion. The target site is at the upper limit of length (25 bases) generated by the endonucleases that catalyse insertion reactions, and all target site-containing sequences are of sufficient length that they can be checked directly by BLASTN. The right duplicated target site has undergone deletions in some Old World monkeys (baboon, macaque) and New World monkeys. As with other EBLN genes (below), the genomic location (coordinates) of EBLN1 is from Kawasaki et al. (2021) or GeneCards (https://www.genecards.org/); the human sequence from the UCSC Browser (https://genome.ucsc.edu/cgi-bin/hgGateway), and that of other species from NCBI BLAST or BLASTN (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Molecular biological research has described functions of the EBLN1 protein in human cells.[24] The protein acts to suppress the accumulation of DNA damage that occurs either spontaneously or following treatment of cells with ionising radiation (a mutagenic agent) and it provides some protection against the lethal effects of radiation on cells. The EBLN1 protein appears to be involved also in regulating the system of microtubules that act to control cell shape, the movement of intracellular components, and the separation of chromosomes when cells divide.
Proteins produced by infectious viruses act to hijack cell functions so as to produce more viruses—typically to the detriment of cells and the organisms comprised of those cells. The equivalent protein found in a contemporary infectious bornaviruses (borna disease virus nucleoprotein, BDV N) does not perform the functions that have been documented for EBLN1. The ability of EBLN1 in human cells to promote cell viability represents a gain of function. It has been coopted into a new role and acquired cell-sustaining properties. It is likely that more molecular research will be done to catalogue how the protein has changed structurally during its residence time in primate organisms.
The insertion site of the second bornavirus-derived gene, EBLN2, is depicted in Figure 4. Once again, the pre-insertion (uninterrupted) target site is apparent in prosimians (tarsier, galago, lemur) and in a variety of non-primate mammals. And, as in the first case, the EBLN2 bornavirus gene is present in New World monkeys (exemplified by the marmoset), Old World monkeys (baboon and macaque), and the apes. It was inserted into the genome of a simian ancestor.
In addition, a transposable element called an Alu element is located immediately to the right of the bornavirus insert (green text; Figure 4). Both components of the insert lie between the duplications of the target site and must have been spliced into the chromosomal DNA at the same time. One can hypothesise that the Alu element recruited the reverse transcriptase enzyme that generated the composite bornavirus-Alu insertion event. Such is a historical reconstruction of a unique molecular event that occurred at least forty million years ago. But the postulated series of events is plausible because reverse transcriptases produced by genomic parasites are still modifying our DNA and are studied in defined molecular biological systems, as noted above (n. 22).

Figure 4. Insertion site of the EBLN2 gene. DNA sequences are shown for nine simian species possessing the insert, and for three prosimian and five non-primate species in which the undisturbed target site could be identified. The first six bases of the bornavirus insert (GGAACC…) are indicated in orange and the first bases of the Alu element (GTCCA…) in green. Human genome coordinates are given in Fujino et al. (2021).
Molecular biological research has described the function of the EBLN2 protein in human cells.[25] It has acquired the ability to localise to mitochondria, organelles that provide energy to drive cell metabolism. (The BDV N protein of contemporary infectious bornaviruses does not localise to mitochondria.) But mitochondria also control life-and-death decisions in cells, and the EBLN2 protein acts to suppress cell suicide. Like EBLN1, this protein has acquired a pro-life role, but it acts by a different mechanism.
The possible activity of one other endogenous bornavirus gene has been investigated. EBLN3P was spliced into the primate germline in the same era as were EBLN1 and EBLN2 (Figure 2), and by the same mechanism (Figure 1; target site duplications are well preserved, Figure 5). However, the EBLN3P sequence does not specify production of a protein; it is known as a pseudogene (as indicated by the P in its symbol). The EBLN3P pseudogene is not totally inert. It is copied into RNA which is expressed at relatively high levels in some breast cancers (of the luminal B subtype). The presence of EBLN3P transcripts is associated with improved prognosis and with less invasive behaviour.[26] The reason for this association is not known. Furthermore, it has been proposed that some EBLN3P-derived sequences have been coopted to generate small RNA molecules (called piRNAs) that protect germline cells from invasion by infectious bornaviruses.[27] Perhaps piRNAs derived from the EBLN3P pseudogene act to protect male fertility.[28]

Figure 5. Insertion site of the EBLN3P pseudogene. In humans, a large deletion (2377 bases) exists immediately to the left of the left-hand target site duplication.
We may summarise the scientific findings hitherto. Millions of years ago, infections with potentially pathogenic bornaviruses scattered foreign bits of DNA through the genomes of our ancestors. Some of these random DNA-modifying events occurred in individuals which would prove to be the ancestors of all simian primates. In at least two cases, heritable (endogenised) inserted bornavirus genes retained the ability to produce proteins and in time acquired new functions. They were integrated into regulatory networks controlling life-and-death decisions in cells. Foreign genes have serendipitously made significant contributions to the biology and survival of contemporary Homo sapiens.
Humanity: Evolutionary History and Divine Creation
This snapshot of bornavirus contributions to our genome and of our “becoming” as humans invites theological interpretation. Many millions of years of our genetic history are minutely documented by the sequence of bases inscribed in our DNA, the genetic text we have inherited. Each randomly added insert has its own history (as illustrated in the cameos of Figures 3 to 5). The acquisition through evolution of functional capacities (mediated in this case by proteins and RNA molecules of bornaviral provenance) is a historical process that can be reconstructed in some detail. Christians believe that God is the sustainer of all histories, including those of biology. God has conferred upon matter the capacity to develop into organic and relational beings of extraordinary capacities. Christians should take with great seriousness what our genome tells us about our evolutionary history. More than that, it directs our worship to its divine originator and sustainer.
Creatio ex vetere: Creation from the Old
In scientific terms, humankind has been generated by a historical process, the mechanisms of which are shared with myriad other creatures and appear to be wholly unexceptional. In theological terms, the advent of humanity represents something qualitatively new, so that humanity is said to be God’s creation,[29] not only in the sense in which all creatures are given being, but also in the sense that humanity represents a striking innovation in the tree of life that is the object of God’s moral address. Our personal capacities are genuinely exceptional. Our createdness confers upon us an inalienable dignity.
If we accept Walton’s proposal that the term creation pertains to the conferring of new function,[30] then we can posit that, from a theological point of view, humanity has been created to fulfil the task of caring for God’s earth and of exercising the privilege of worshiping the God to whom the world owes its existence.[31] The advent of the new entity of humanness from a long evolutionary past could be described as creatio ex vetere. This is creation from the old, creation from preexisting matter and from progenitor creatures (including viruses!) that lacked the more fully developed features and responsibilities that define Homo credens, believing humanity, the species that makes metaphysical commitments.
The creation of humanity represents a new reality in the progressive sequence of God’s originating works. Another manifestation of creatio ex vetere is the incarnation of the divine Son, for whom a body was prepared[32] in the divine initiative that constituted the dawn of the new creation. As Adrio Konig emphasised, “the incarnation is an event of decisively eschatological character.”[33] In Jesus of Nazareth, God’s self-revealing Word, God lived among sinful and oppressed humanity as fully as God will live among redeemed humanity in the completed creation.[34] In Jesus we see the presence of the eternal in time.[35] To be fully human, Jesus’ body, like ours, would have been that of an anthropoid primate, complete with its retroviral and bornaviral contributions. It was this evolved body in which “the full content of divine nature lives.”[36] It was this flesh-and-blood human nature that qualified him to be the high priest of mortal humanity.[37]
God’s creatio ex vetere also pertains to the resurrection of Jesus, in which his mortal body was raised transformatively to the unprecedented state of immortality.[38] Resurrection itself represents the inauguration of a new world, a new creation: “A new world has dawned in which forgiveness of sins is not simply a private experience; it is a fact about the cosmos.”[39] Human beings in their inherent territoriality and selfishness will be redeemed by Christ and transformed into a new community that bears the character of Jesus and is at home in the ecology of a new creation. Resurrection is itself a paradigmatic manifestation of the transformation of the old creation into the new. We are presented with “the already existing reality of new creation from within the old.”[40] As Tom Wright states of creatio ex vetere: “The point of new creation is that it is the redemption and transformation of the present creation.”[41]
Creation Involves the Conferral of God’s Image and Likeness
The concept of creatio ex vetere mirrors the idea, pervasive in Scripture, of our being created in “God’s image and likeness.” This term carries three different referents, reflecting the climaxes of three phases of history.
First, our possession of God’s image and likeness denotes our common humanity,[42] the embodied product of millions of years of evolution (including forty million years of our three endogenous bornavirus contributions) as described in what I call the “Primal Testament,” the genome. Van Huyssteen summarises the concept of the imago Dei as embodied human uniqueness.[43] But when was this status acquired? Scientific humanity would like to know “when and how humans were created in God’s image.”[44]
This term does not refer to any one feature we possess, such as rationality, creativity, moral sense or the capacity for relationality. Rather, the image of God is said to describe our functional status;[45] it refers to our calling by God, our vocation,[46] our commissioning as God’s agents on earth. To Briggs et al., it pertains to accountability or responsibility to God.[47]
The personal properties required to be God’s representatives required an evolved neural substrate and the cultural underpinnings that developed over no less than 200,000 years, the age of anatomically modern Homo sapiens. But these were merely the prerequisite capacities needed to engage with God in the personal dimension featuring relationship and obedient service.
Humans could be said to possess God’s image only when called into service as God’s representatives on earth, which presupposes that they could (at least potentially) respond.[48] In this case, the concept of the imago Dei would arise from that phase of the missio Dei when God addressed human beings. The human vocation to serve God in creation is coeval with God’s mission to redeem a frustrated, painfully incomplete, and suffering creation,[49] and with Israel’s awareness that God’s call is directed equally to all people, not merely the elite (as in pagan Mesopotamian thought).[50] Israel’s horizons of the imago Dei encompassed the humanity Israel actually knew and was called to serve. The perspective of humanity’s call should define our self-understanding, even as we have come to appreciate more the challenge of our biological (including viral) and prehistorical antecedents.[51]
A second use of the divine image pertains to the denouement of a second history—that of Abraham’s family as described in the Old Testament. This history also was marked by contingency—often misused freedom, moral failure, and cataclysmic judgment. But this history was also resoundingly fruitful in that it climaxed in the advent of Jesus, who was the image of God,[52] and specifically the express or exact image and likeness of God.[53] Jesus was the perfect representation of all that humanity and Israel were intended (but failed) to fulfil. In Jesus had come at last “a truly human being … whose aim was to rehumanize other humans … and to re-establish them as what they were supposed to be.”[54] Once again, the messiness of history, this time more particularly Israel’s falteringly human one, has issued in a glorious advance in God’s plan for his creation.
The third climax of history is described in the New Testament. The history of Jesus and his church will culminate in the conferment of the perfect image and likeness of God, as present in Jesus, upon the earthy creatures who had so faithlessly represented God hitherto: “Just as we wear the likeness of the man made of earth, so we will wear the likeness of the Man from heaven.”[55] With this transformation, the earthiness of the sinful hominoid primate receives the nature of the incarnate Son of God. The new humanity will be consummately created from the old.
Potential for Evil and Good
Biological (including genetic) history witnesses to the actions of agents that exert ambiguous effects. In the short term, agents such as retroviruses and transposable elements (which acted to endogenise bornavirus RNA) are mutagens, genome disruptors, potential pathogens, and typically accumulate in the genome as junk (they are degenerative). In the long term, they can be recruited to provide essential functions, both structural and regulatory (they are generative), and they confer evolvability upon their host organisms. Bornaviruses themselves are pathogens, but have contributed to our genetic endowment. God is responsible for biological and human histories that are replete both with terrible suffering and inspiring beauty. Is God culpable for the suffering?
The potential in God’s creation for good and evil is manifested also in the scientific enterprise itself. While writing this paper, I perused several publications purporting to show that an endogenous bornavirus gene performed multiple regulatory functions. However, the papers contained anomalies that cast doubt on their veracity.[56] Fraudulent publications pervade the biomedical literature.[57] If science is God’s creation, a gift of God,[58] then it is inherently good. However, the history of science, like other created histories, is ambivalent. This ambiguity arises because people may exercise their freedom to act either in ways compatible with God’s wisdom (truthfully, such that science flourishes) or that contravene God’s wisdom (dishonestly, such that science withers). Creation is good but the creatures (impersonal matter or personal agents, including those who would exploit science for their own nefarious ends) are free. It is necessarily free human agents, not God, who are culpable for duplicitous actions. Similarly, it is free process that has beautiful or harmful outcomes in God’s good world.
This pattern is inherent to biblical history. Israel’s history in the short term appeared to be a random mess, in which God’s laws and spokespeople were often rejected. Israel’s history seemed to end in disaster. But from the perspective of the New Testament, in the long term there was forward movement, the anticipation of deliverance, and the great culmination of God’s Messiah as the paradigmatic human being, the yearned-for climax of Israel’s history.[59] Jesus’ own mission seemed to have been a failure—he was controverted, rejected, betrayed, crucified—but was ultimately vindicated by resurrection, which none of his followers had remotely anticipated.
Evil precedes and may be the substrate out of which good arises. The happenstance of biological evolution with its concomitant costs and gains finds a parallel with the biblical motif of suffering and glory. John’s gospel takes the “suffering and glory” theme back to Jesus himself: “‘That is why I came—so that I might go through this hour of suffering. Father, bring glory to your name.’ Then a voice spoke from heaven, ‘I have brought glory to it, and I will do so again.’”[60] Luke quotes Jesus similarly: “Was it not necessary for the Messiah to suffer these things and then to enter his glory?”[61] And other New Testament writers recognise its validity. Paul writes: “I consider that what we suffer at this present time cannot be compared at all with the glory that will be revealed to us.”[62] We cannot separate cross and kingdom motifs in the gospel of Jesus[63] any more than we can separate them in biological history.
But what about the sacrifice of numerous individuals (especially children) who have suffered genetic disease and cancers and the depredations of evolved pathogens as a result of the same processes that have led to the advent of the wonders of life and of humanity? Science suggests that the only possible world is one in which randomness and freedom operate. Polkinghorne has said that suffering and evil are the “inescapable cost” of a creation “permitted to be itself.” He stated that “the possibility of cancer is the necessary price of the evolution of new life.”[64] Christian cosmologist Heino Falcke has described how solar cosmic radiation is both a driver of evolution and a source of cancers. Our existence as human beings “has been earned at the cost of deep suffering. But without these potentially dangerous genetic changes, we would still be single-celled organisms.”[65] As McLeish has noted, if we are to eliminate randomness by reducing the temperature to absolute zero, we necessarily eliminate life too.[66]
It may be argued logically that “the existence of good” requires “the possibility of evil.” That is cold comfort. But our capacity to endure suffering is best sustained by the unconquerable divine love demonstrated in Jesus’ suffering on the cross.[67] As Polkinghorne said, “God is a fellow participant in the world’s suffering … This is one of the meanings of the cross of Christ.”[68] Tom Wright has likewise said that the suffering of Jesus (to which his followers are called) does not merely accompany the attainment of God’s purposes but is the necessary means by which they are achieved.[69] And similarly, for Jesus’ followers, suffering is not merely something to be endured; it also has the “positive effect of carrying forward the redemptive effect of Jesus’ own death … by sharing in it.”[70] Suffering is more than a dark tunnel to be traversed en route to the Kingdom of God; it is the effective way of achieving the goal.[71] Questions posed currently by biological history have been addressed by salvation history.
God has compassion over all he has made.[72] Brueggemann states that “the giver of abundant life generates a world of blessing where none seemed possible.” God brings life and fruitfulness out of situations within which chaos and barrenness seem to prevail, transforming “scenes of hopelessness into occasions of life, possibility, and joy.”[73] When the randomness inherent to life leads to intolerable grief, people can only trust that God is just, and that God suffers with his creatures redemptively. Genuine compassion for fellow-creatures enduring afflictions that are concomitant with an evolving world should also be manifested in acts of selfless charity.
We need to be reminded of what Charles Raven wrote in 1955, when he provided a theological interpretation of the evolutionary paradigm:
It is one of the ironies of history that Christendom which by its own Scriptures was committed to belief in an ever-working God (e.g. John 5:17) in a progressive revelation still incomplete (John 16:13), in suffering as the characteristic of the creature (Rom. 8:18-23) and the means to perfection (Hebr. 2:10), and in fuller life as the divine purpose (John 10:10) should have so signally failed to maintain this belief when faced with the challenge of Darwinism.[74]
Hubris or Humility
Our genome is an eclectic hodgepodge of DNA from multiple sources. It seems that our lives are enriched, if not sustained, by genes contributed by potentially pathogenic viruses. Our descent from ancestors we share with monkeys is assured. Some might consider that such claims are an insult to the creator whose image and likeness we bear; and that we demean ourselves, the crown of creation.
Our heritage of viral componentry does indeed emphasise our humble origins. There is no room for hubris. But this is all part of the way by which God’s ends are achieved in history. Israel was told: “The LORD did not set his affection on you and choose you because you were more numerous than other peoples, for you were the fewest of all peoples.”[75] Human societies cannot abide thoughts of their own insignificance. National histories glorify their own past. Some sort of jingoism underlies tribal and national self-evaluation—hence the horrors of tribalism and nationalism. In contrast, in the (perhaps unique) case of Israel, “biblical history constantly confesses their failure as a renegade people, and glorifies the God who made something of these historical nobodies in spite of themselves and their repeated disobedience.”[76]
Similarly, the first followers of Jesus in the nascent church were reminded that few of them “were wise or powerful or of high social standing,” which meant that no one could boast in God’s presence.[77] The church was composed of people who were spiritually dead but brought to life in Christ.[78] The great figures of Israel’s and the church’s story were in themselves deeply flawed. When called by God, Moses confessed to being too halting; Isaiah, too impure; Jeremiah, too young; Peter too compromised by fear; Paul, too hostile.[79]
As noted above, we are composed of earth, humans from humus, vitality (in part) from viruses, even virulent ones. To be told that I have an ape as an ancestor on my mother’s side (a point of contention in the famous Huxley-Wilberforce debate) is a mild put-down by comparison with the discovery that I am part virus. As a result of our heritage of viral flotsam, materialists may see themselves as inconsequential cosmic accidents. The late E. O. Wilson asserted that “Darwin showed that humanity is not the centre of creation, and not its purpose either.”[80] But, as Gingerich observes, Darwin the scientist could not have shown this. Wilson’s proposal is merely a feature of his philosophical stance or ideology. Our inestimable value is conferred upon us by God, who calls us into the service of the Kingdom of God. With Wright, Christians believe that Jesus is the one in whom God “has acted in cosmic history, human history, and Israel’s history to do for Israel, humanity and the world what they could not do for themselves.”[81] The whole physical universe, including its living organisms and their genetics inscribed in DNA, comes to fulfilment only in Jesus. Indeed, the totality of history, “all space, time and matter was summed up in this king.”[82] Our value then, comes not from the raw material of which we are constituted, but from what God intends to do with it.
To conclude, scientific (genetic) research has shown that segments of bornaviral genomes have been inserted randomly into the genomes of animals. Some of these viral genes acquire new functions in the host organisms. Virus-derived genes are part of our own genetic heritage. A theological interpretation perceives that such happenstance fits into a pattern, observed in biblical history—and indeed our personal histories—by which God transforms the old into the new (creatio ex vetere), randomness into meaningfulness, suffering into glory. Bornaviruses alert us to the earthiness of our biological origins, and to our place in a cosmic history that is both free and directed, and by which God’s purposes will be realised.
The author reports there are no competing interests to declare.
Received: 17/05/23 Accepted: 21/07/23 Published: 09/08/23
[1] Graeme Finlay, “Evolution as History: Phylogenetics of Genomes and Manuscripts,” Christian Perspectives on Science and Technology, New Series 1 (2022): 150–174, https://doi.org/10.58913/JJHH2131.
[2] Graeme Finlay, “Being and Becoming: The Complementarity of Creation and Evolution,” Christian Perspectives on Science and Technology, New Series 2 (2023): 1–27, https://doi.org/10.58913/RDDN1562.
[3] David M. Morens and Anthony S. Fauci, “Emerging Pandemic Diseases: How We Got to COVID-19,” Cell 182 (2020): 1077–1092, DOI: 10.1016/j.cell.2020.08.021.
[4] Cristina Frank et al., “Human Borna Disease Virus 1 (BoDV-1): Encephalitis Cases in the North and East of Germany,” Emerging Microbes and Infections 11 (2022): 6–13, DOI: 10.1080/22221751.2021.2007737; Monika Huhndorf et al., “Magnetic Resonance Imaging of Human Variegated Squirrel Bornavirus 1 (VSBV-1) Encephalitis Reveals Diagnostic Pattern Indistinguishable from Borna Disease virus 1 (BoDV-1) Encephalitis but Typical for Bornaviruses,” Emerging Microbes and Infections 12 (2023): 2179348, DOI: 10.1080/22221751.2023.2179348. It appears that the virus enters by the nose and is transmitted via olfactory neurons to deep parts of the brain.
[5] For an overview see Masayuki Horie, “The Biological Significance of Bornavirus-Derived Genes in Mammals,” Current Opinion in Virology 25 (2017): 1–6, DOI: 10.1016/j.coviro.2017.06.004.
[6] Masayuki Horie and Keizo Tomonaga, “Paleovirology of Bornaviruses: What Can be Learned from Molecular Fossils of Bornaviruses,” Virus Research 262 (2019): 2–9, DOI: 10.1016/j.virusres.2018.04.006.
[7] Junna Kawasaki et al., “100–My History of Bornavirus Infections Hidden in Vertebrate Genomes,” Proceedings of the National Academy of Sciences of the USA 118 (2021): e2026235118, DOI: 10.1073/pnas.2026235118.
[8] For the sake of focus, in this paper only three insertion mutations (Figures 3–5) have been described in detail. Millions of inserted transposable elements are shared by humans with other primates. For a study of the few (exceptional) human-specific transposable elements, see Maria V. Suntsova and Anton A. Buzdin, “Differences between Human and Chimpanzee Genomes and their Implications in Gene Expression, Protein Functions and Biochemical Properties of the Two Species,” BMC Genomics 21 (2020): 535, DOI: 10.1186/s12864-020-06962-8. At greater phylogenetic distances, humans share many thousands of transposable element insertions with non-primate mammals, and hundreds of more ancient transposable elements with non-mammal vertebrates. A recent study has documented 882 inserts that entered our DNA in amniote ancestors (of mammals and reptiles, as well as birds), 35 in tetrapod ancestors (of amniotes and amphibians) and eight in gnathostome ancestors (of tetrapods and fish, including sharks). See Martin C. Frith, “Paleozoic Protein Fossils Illuminate the Evolution of Vertebrate Genomes and Transposable Elements,” Molecular Biology and Evolution 39 (2022): msac068, DOI: 10.1093/molbev/msac068.
[9] Genesis 2:7; Psalm 90:3.
[10] Jude 8; cf. Hebrews 9:13; J. D. Douglas et al. (eds), The Illustrated Bible Dictionary (Leicester: IVP, 1980), 510.
[11] Matthew 26:41; Mark 14:38.
[12] 2 Corinthians 7:5; translated as “body,” NIV.
[13] Galatians 5:19–21, 24; Romans 8:5–6; Ephesians 2:3; translated as “human nature” or “natural desires,” GNT; “sinful nature,” NIV; “lower nature” or “carnal attitude,” Phillips.
[14] Galatians 5:17; Romans 8:7–8.
[15] N. T. Wright, Paul and the Faithfulness of God (London: SPCK, 2013), 1020.
[16] John 6:63; translated as “human power,” GNT.
[17] John 1:14; 1 John 4:2.
[18] John 6:51–56; Ephesians 2:14; 1 Peter 3:18.
[19] Galatians 2:20; Ephesians 3:19; 2 Pet 1:4.
[20]. Graeme Finlay, Evolution and Eschatology: Genetic Science and the Goodness of God (Eugene, OR: Wipf and Stock, 2021), 164–168.
[21] Finlay, “Evolution as History,” 150–174.
[22] Endonuclease and reverse transcriptase enzymes encoded by transposable elements and endogenous retroviruses have been characterised in detail biochemically and structurally. See Ian Miller et al., “Structural Dissection of Sequence Recognition and Catalytic Mechanism of Human LINE-1 Endonuclease,” Nucleic Acids Research 49 (2021): 11350–66, DOI: 10.1093/nar/gkab826; Eric T. Baldwin et al., “Human Endogenous Retrovirus-K (HERV-K) Reverse Transcriptase (RT) Structure and Biochemistry Reveals Remarkable Similarities to HIV-1 RT and Opportunities for HERV-K-Specific Inhibition,” Proceedings of the National Academy of Sciences of the USA 119 (2022): e2200260119, DOI: 10.1073/pnas.2200260119.
[23] Liliya Doronina et al., “True Homoplasy of Retrotransposon Insertions in Primates,” Systematic Biology 68 (2019): 482–493, DOI: 10.1093/sysbio/syy076.
[24] Katie N. Myers et al., “The Bornavirus-Derived Human Protein EBLN1 Promotes Efficient Cell Cycle Transit, Microtubule Organization, and Genome Stability,” Scientific Reports 6 (2016): 35548, DOI: 10.1038/srep35548.
[25] Kan Fujino et al., “A Human Endogenous Bornavirus-Like Nucleoprotein Encodes a Mitochondrial Protein Associated with Cell Viability,” Journal of Virology 95 (2021): e02030–20, DOI: 10.1128/JVI.02030-20.
[26] Carolina Mathias et al., “Unraveling Immune-Related lncRNAs in Breast Cancer Molecular Subtypes,” Frontiers in Oncology 11 (2021): 692170, DOI: 10.3389/fonc.2021.692170.
[27] Hirohito Ogawa and Tomoyuki Honda, “Viral Sequences Are Repurposed for Controlling Antiviral Responses as Non-Retroviral Endogenous Viral Elements,” Acta Medica Okayama 76 (2022): 503–510, DOI: 10.18926/AMO/64025; citing Nicholas F. Parrish et al., “piRNAs Derived from Ancient Viral Processed Pseudogenes as Transgenerational Sequence-Specific Immune Memory in Mammals,” RNA 21 (2015): 1691–1703, DOI: 10.1261/rna.052092.115.
[28] Tomoko Takahashi, Steven M. Heaton, and Nicholas F. Parrish, “Mammalian Antiviral Systems Directed by Small RNA,” PLoS Pathogens 17 (2021): e1010091, DOI: 10.1371/journal.ppat.1010091.
[29] Genesis 1:27; 5:1–2; 6:7; Deuteronomy 4:32; Psalm 89:47; 102:18; Isaiah 45:12, as listed by John H. Walton, The Lost World of Genesis One (Downers Grove: IVP, 2009), 41–44.
[30] Walton, Lost World, 54–71.
[31] Walton, Lost World, 68, sees “image of God” as a “functional element” of Genesis 1:26–30; see below.
[32] Hebrews 10:5; and which, as Dr Murray Harris once stated (personal communication), the church should celebrate on Annunciation Day, 25 March, not Christmas.
[33] Adrio Konig, The Eclipse of Christ in Eschatology (Blackwood, South Australia: New Creation Publications, 2007; first edn 1989), 69.
[34] Konig, Eclipse, 71; the divine Word Jesus has dwelt (ἐσκήνωσεν; Jn 1:14) and God will dwell (σκηνώσει; Rev 21:3) among us.
[35] M. Michaelis in Konig, Eclipse, 72.
[36] Colossians 2:9 GNT.
[37] Hebrews 2:14–18.
[38] 1 Corinthians 15:42–56.
[39] N. T. Wright, Surprised by Hope (New York: HarperOne, 2008), 246–247.
[40] N. T. Wright, Surprised by Scripture (London: SPCK, 2014), 203.
[41] Wright, Surprised by Scripture, 201.
[42] Genesis 1:26–27; 9:6; James 3:9.
[43] J. Wentzell van Huyssteen, Alone in the World? (Grand Rapids: Eerdmans, 2006), 159–163. Scholars cited provide more specific meanings of the imago Dei: “a specificity gained from being addressed by God’s moral word, and the ability to respond, especially in prayer” (Robert Jenson); “that which [in humans] portrays or sets forth God in the world” (Philip Heffner); a term that indicates both an analogy between God and humans (is representational) and the caring task entrusted to humanity by God (is representative) (Richard Middleton); 145–149, 156–158, 273–274.
[44] Ian Hore-Lacy, review of The Faraday Papers, https://journal.iscast.org/book-reviews/review-the-faraday-papers.
[45] Paul Copan and Douglas Jacoby, Origins: The Ancient Impact and Modern Implications of Genesis 1–11 (New York: Morgan James, 2019), 56–58. We further read, “As God’s image, we represent him and join him in his kingdom work” (64).
[46] Wright, Surprised by Scripture, 35, 159.
[47] Andrew Briggs, Hans Halvorson, and Andrew Steane, It Keeps Me Seeking (Oxford: Oxford University Press, 2018), 74.
[48] Given that our possession of God’s image is an act of grace, we should allow that that same grace is extended to those who, for whatever reason (age, accident, genetics) have a diminished capacity to respond to God. In such cases, we should be content to acknowledge that, as God is a mystery, so is God’s image a mystery. See Janet Martin Soskice, “Imago Dei and Sexual Difference: Toward an Eschatological Anthropology” in Rethinking Human Nature: A Multidisciplinary Approach, ed. Malcolm Jeeves (Grand Rapids: Eerdmans, 2011), 295–308, esp. 297, 325. The reality of the divine image in us is evinced by the compassion and care we show to persons with disabilities.
[49] Romans 8:20.
[50] Denis Alexander, Are We Slaves to Our Genes? (Cambridge: Cambridge University Press, 2020), 196–214.
[51] Our constitution as earth, our vitality as divine breath (Genesis 2:7), our calling as image (Genesis 1:26–27), and our status before God as disobedient (Genesis 3) all describe the universal human condition. They are theological anthropology, rather than discrete events in the past. They are not physical anthropology.
[52] Colossians 1:15.
[53] 2 Corinthians 4:4; Hebrews 1:3.
[54] Wright, Paul, 377; also 406.
[55] 1 Corinthians 15:49; also 2 Corinthians 3:18; Romans 8:29; Ephesians 4:24; Colossians 3:10–11; 1 John 3:2.
[56] I have corresponded with editors of several journals, and at the time of writing await their assessment.
[57] Jennifer A. Byrne et al., “Protection of the Human Gene Research Literature from Contract Cheating Organizations Known as Research Paper Mills,” Nucleic Acids Research 50 (2022): 12058–12070, DOI: 10.1093/nar/gkac1139. The frequency of fraudulent papers in medicine could be as high as 24%. See Jeffrey Brainard, “New Tools Show Promise for Tackling Paper Mills,” Science 380 (2023): 568–569.
[58] Graeme Finlay, God’s Gift of Science (Eugene, OR: Wipf and Stock, 2022); David Hutchings and Tom McLeish, Let There be Science: Why God Loves Science, and Science Needs God (Oxford: Lion Hudson, 2017), 172, 178, 184, 188. Upon rereading Hutchings’ and McLeish’s book, I have wondered whether I subconsciously used as a title for my book a term they developed. If so, I belatedly acknowledge my indebtedness to them.
[59] Romans 9:5.
[60] John 12:27–28.
[61] Luke 24:26; described by Tom Wright as the “cross and kingdom” motif, in How God Became King (London: SPCK, 2012), 183–184; 139, chs 9, 10.
[62] Romans 8:18; Hebrews 2:9; and the preaching (Acts 3:18 [suffering], 13 [glory]) and writing of Peter (1 Peter 1:7; 1:11; 4:13; 5:1; 5:10).
[63] Wright, How God Became King, 159–160.
[64] John Polkinghorne, Quarks, Chaos & Christianity (London: SPCK, 1994), 47–48.
[65] Heino Falcke, Light in the Darkness (London: Wildfire, 2021), 25–26.
[66] Tom McLeish, “Evolution as an Unwrapping of the Gift of Freedom,” Scientia et Fides 8 (2020): 43–64, DOI: 10.12775/SetF.2020.014.
[67] Robert F. L. Boyd, “The Space Sciences,” in Horizons of Science, ed. Carl F. H. Henry (New York: Harper and Row, 1977), 1–20; John Houghton, The Search for God (Oxford: Lion, 1996), 188.
[68] Polkinghorne, Quarks, 48.
[69] Wright, How God Became King, 199.
[70] Wright, How God Became King, 201.
[71] Wright, How God Became King, 237.
[72] Walter Brueggemann, Theology of the Old Testament (Minneapolis: Fortress, 1997), 218 (citing Ps 145:8–9).
[73] Brueggemann, Theology, 204–205, 207.
[74] Charles E. Raven, Christianity and Science (London: Lutterworth, 1955), 31.
[75] Deuteronomy 7:7.
[76] Harold Turner, The Roots of Science (Auckland: DeepSight Trust, 1998), 78.
[77] 1 Corinthians 1:26.
[78] Ephesians 2:1–9.
[79] Exodus 4:10; Isaiah 6:5; Jeremiah 1:6; Mark 14:71–72; 1 Corinthians 15:9; Ephesians 3:8; 1 Timothy 1:15–16.
[80] In Owen Gingerich, God’s Universe (Cambridge, MA: Harvard University Press, 2006), 98. For Wilson to raise issues like “centre of creation” and “purpose,” he wanders into metaphysics. He is not speaking as a scientist. We must be alert to such covert “religious” talk by people purporting to represent science.
[81] Wright, Paul, 684.
[82] Wright, Paul, 731.