
Christian Perspectives on Science and Technology, New Series, Vol. 2 (2023), 1–27
Abstract: Longstanding debates relating to biological evolution concern whether random events (mutations of DNA) are able to generate new functionality, and whether such proposed evolutionary mechanisms are compatible with belief in divine creation. The sequencing of genomes from multiple species has generated a flood of genomic data, so that genetic changes may be correlated with species’ phenotypes. Our genomes are modified by mutagenic agents such as retroviruses (ERVs) and transposable elements (TEs). Empirical data confirm that random accumulations of ERVs and TEs in the human genome have rewired regulatory networks in early embryos (ERV-like MaLR elements), embryonic stem cells (ERV-H), and primordial germ cells (ERV-K). Altered regulation of gene activity in neural cells has been evinced for a class of TEs called SVA elements. Random, stochastic events in the context of natural laws that are hospitable to life may indeed generate new genetic information. Christians may see such phenomena as aspects of a freely operating and fruitful creation. Acceptance of biological evolution and the role of randomness in an anthropic cosmos are indeed compatible with the biblical concept of creation—that the whole system is ordained, ordered, and sustained by a purposeful and self-revealing God.
Of longstanding interest to biology is the question of whether random mutations are able to generate new forms and functions during evolution. This issue has been likened to the question of whether myriad monkeys pecking away randomly at typewriters for a very long time could generate the works of Shakespeare. Or whether a tornado in a junk yard might assemble a Boeing 707.
The development of comparative genomics can now provide empirical results that throw light on this question. The genomes of thousands of species have been sequenced. Genome sequences from different species can be aligned with each other to identify mutations that have appeared in particular taxa of organisms (reflecting particular stages of evolution). Increasingly, mutations can be related to a species’ phenotype to indicate whether random genetic changes can underlie the development of regulatory networks, integrated functions, and complex structures.
For many people who are interested in the question of a creator, these considerations seem to provide the possibility of an answer. It might be thought that if random mutations can account for new features in biological evolution, then a creator is no longer necessary. This paper reviews recent evidence that random mutations—featuring insertions of viral sequences and of transposable elements into genomic DNA—do indeed generate features peculiar to human biology. Random events certainly underlie the acquisition of characteristic features of Homo sapiens.[1]
But these discoveries cannot address the question of a creator. Randomness alone can generate nothing; it requires the context of a lawful anthropic cosmos—that is, a universe with the potential to sustain life such as ours. The fruitful interplay of free randomness and directing lawfulness underlies evolution. The question of a creator pertains to the very existence of such a world. We accept by faith the idea that this productive cosmos is divinely ordained. Or equally, we reject by faith the idea that this fecund cosmic structure comes from the mind of God. (By faith, I intend to indicate commitment in the absence of absolute certainty.) No scientific discovery can adjudicate between these alternatives. Those who perceive an incompatibility between biological evolution and theological creation are misinformed.
It follows that belief in God’s creative activity does not pertain to individual events that are describable by science; but to the very existence of a cosmos in which free and lawful events can occur. Ultimately, of course, belief in a creator is based on personal communication—God’s self-revelation—which Christians believe has occurred in Jesus.
Endogenous Retroviruses
We have discussed in a previous article[2] the fact that most of our genome is composed of randomly accumulated units of parasitic DNA. Endogenous retroviruses (ERVs) and transposable elements (TEs) are semi-autonomous genetic elements that randomly colonise (and so modify) the genomes of (probably) all organisms. (My focus on evolutionary relationships between humans and other placental mammals—for which ERVs and TEs provide unambiguous phylogenetic markers—means that I will not consider more distantly related organisms such as invertebrates or bacteria, or the mutagenic mechanisms that pertain to those organisms.) The way ERVs and TEs proliferate in genomes is stochastic—the time and location at which a new element will arise cannot be predicted—but repeatedly they have been recruited into providing new functions.
New elements arise in the genome as junk—as unsolicited accretions to the genome of a functioning organism—and some of them initiate genetic disease.[3] Disease-causing elements are pathogenic junk. But there is growing evidence that a proportion of ERV and TE insertions eventually acquire functions that serve the host organism. Units of DNA added randomly to genomes may transition from junk to valuable or even essential genetic componentry.
Some ERVs have contributed genes that function in the development of the placenta. ERV genes usually decay with time from insertion into animal genomes. But a small number retain protein-coding capacity, presumably because the viral protein contributes to the survival or reproductive success of the host animal. Of particular interest are retroviral envelope genes, that enable the viruses to adhere to cells during the process of infection. Some ERV envelope genes have been domesticated to specify the production of proteins (now called syncytins) that promote the formation of the syncytiotrophoblast, the lining of the placenta.[4] Another retroviral envelope protein has been transformed into a derivative (now called suppressyn) that may act to regulate or tone down excessive syncytin activity.[5] In addition, suppressyn prevents envelope protein on infectious retrovirus particles from adhering to placental cells. This domesticated retroviral protein acts to provide a defensive barrier that prevents invading retroviruses from docking on to cells.[6] Part of a retrovirus pol gene has been transmogrified into a gene (NYNRIN) that contributes to the invasion of placental trophoblast cells into the uterus.[7]
ERVs have been coopted into roles which rewire or reconfigure genomic control circuits. Such reorganisation may include the specification of the body plan early in embryonic development.[8] Three classes of ERV will be considered below. They appear to exert regulatory influences in stem cells during early phases of ontogeny.
MaLR elements are ERV-like entities that colonised the genomes of (now extinct) primates from which monkeys and apes are descended. Many such MaLR elements in our genome become active very early in embryonic development: at the four-cell and eight-cell stage of our personal histories. They provide binding sites for a protein called DUX4 that is a master regulator of genetic expression. (In humans, but not macaques, some of these MaLR elements are active also in the adult pineal gland and bind the OTX2 protein to regulate other genes.)[9] Randomly accumulated retroviral DNA segments have been recruited to orchestrate our primordial genetic programme.
It has been known for some time that members of a class of ERV (ERV-H) are genetically active—they are copied or transcribed into RNA molecules—in pluripotent embryonic stem cells.[10] Such ERVs are believed to be involved in maintaining stem cell pluripotency. Recent findings have identified a particular subset of ERV-H sequences (LTR7up) that is responsible for the maintenance of pluripotent stem cells. What is common to these functional ERV sequences is that they possess a DNA sequence motif (ACAAAAGA) to which regulatory proteins (SOX2 and SOX3) can bind, and thereby activate nearby genes implicated in stem cell maintenance (as depicted, Figure 1).[11]
The insertion site of one of these ERV-H sequences is depicted in Figure 1. The viral sequences (in green font) start with CAGG… (on the left) and, hundreds of bases later, end with …GCATG on the right. This ERV-H is present at the identical genomic site in all the African great apes. That means that it arose in a unique event that occurred in an ancestor common to these four species. However, the ERV-H sequence is absent, and the target site undisturbed, in Asian apes, Old World monkeys, and New World monkeys. This potentially functional element arose in primate DNA by the standard random infectious mechanism.
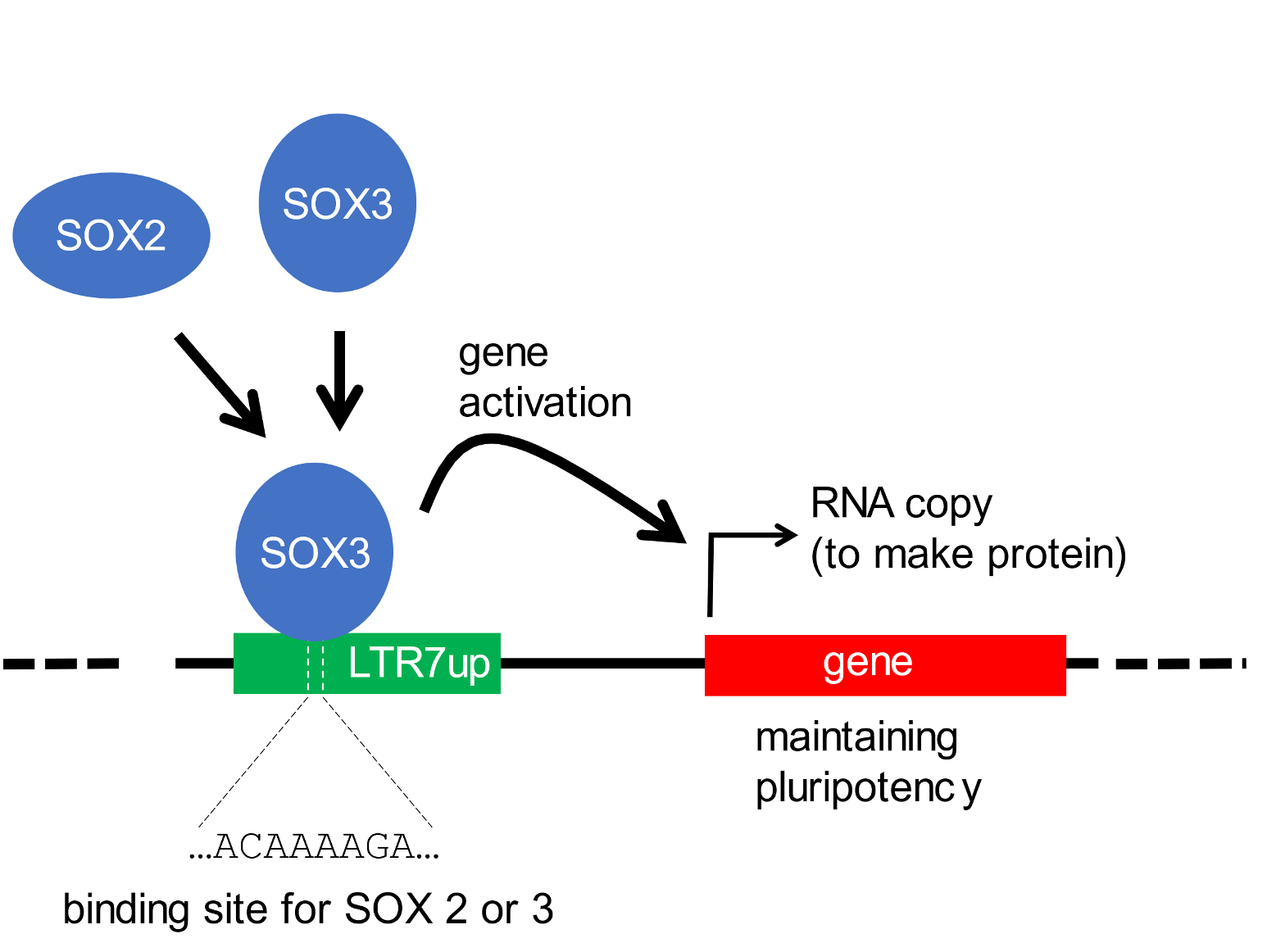
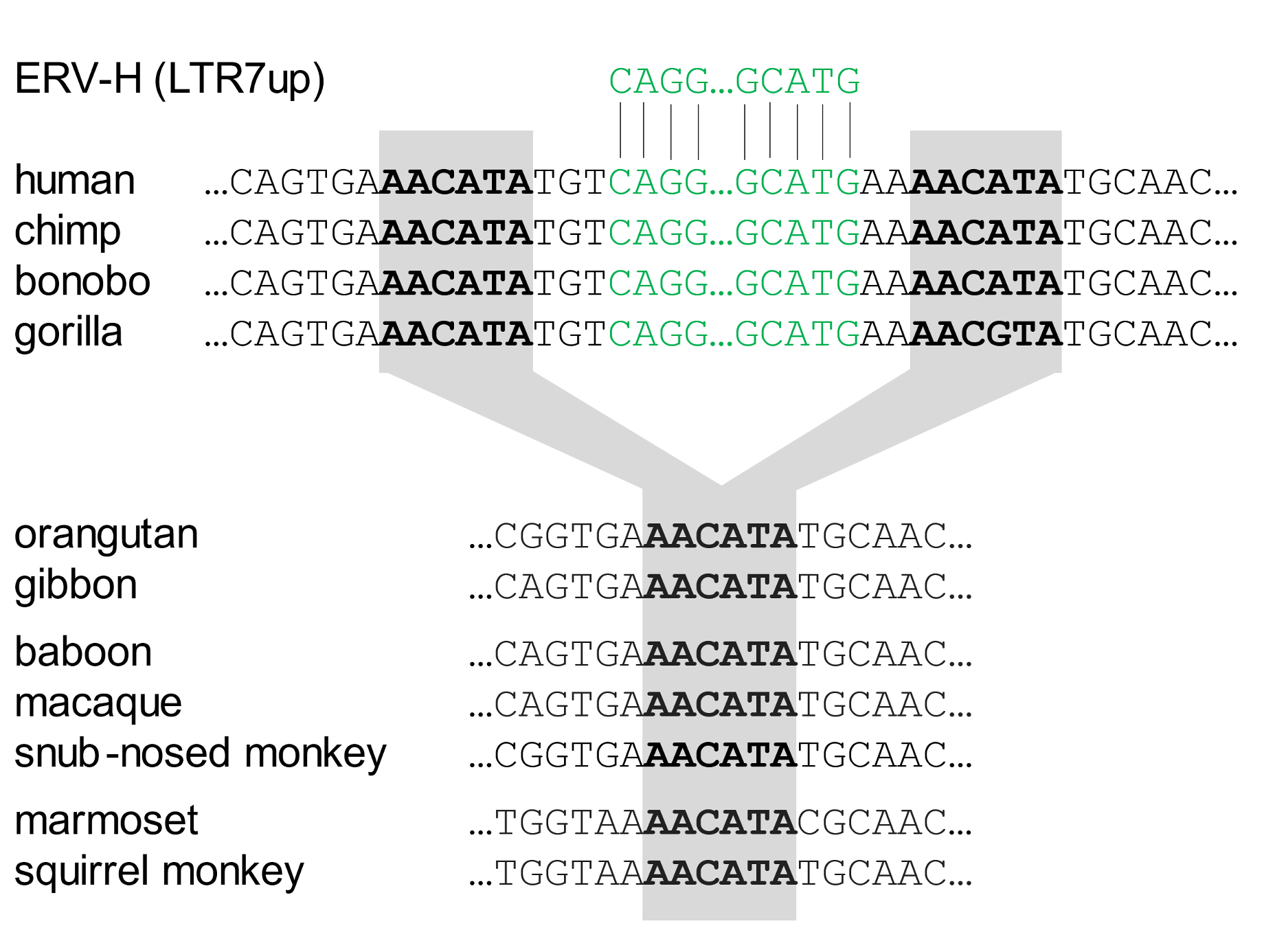
Figure 1. An ERV-H insert (subgroup LTR7up) activates genes in pluripotent stem cells Above, a generalised scheme depicting an LTR7up insert (green box) in which a series of bases (…ACAAAAGA…) recruits proteins (SOX2 or SOX3) that activate a nearby gene (red box) with the function of sustaining pluripotency. Below, an insertion site of an LTR7up element. The ERV sequence is depicted in green font; the target site (AACATA) and its duplications are in bold type and shaded. This insert was from ref. 11, https://elifesciences.org/articles/76257/figures#files, supplementary file 1, row 5. In this and following figures, sequences were recovered using the UCSC Genome Browser and the NCBI BLAST algorithm
A second subclass of endogenous retrovirus, ERV-K (subclass LTR5Hs), has been recruited into gene-regulating circuitry early in embryonic development. ERV-K inserts are genetically active—they are transcribed into RNA—in primitive pluripotent stem cells and also in primordial germ cells. The latter cells are a class of stem cell that are produced in the early embryo and that give rise to reproductive cells (eggs and sperm). Primordial germ cells are vital for fertility.[12]
One particular integrant is found near the FHAD1 gene and its exact point of insertion is depicted in Figure 2. As with Figure 1, the insertion event occurred in an African great ape ancestor, and the undisturbed pre-integration site is present in orangutan and gibbon, monkeys, prosimians, and even some non-primate mammals.
In humans, as pluripotent stem cells transform into primordial germ cells, ERV-H activity decreases and ERV-K activity increases. It is intriguing to consider that randomly accumulated ERV types coordinate the activities of differing genetic programmes as cells progress through sequential stages of development. Retroviruses long resident in our DNA control the early stages of our development as human beings and, according to the most recent evidence, they continue to do so as the body matures.[13]

Figure 2. An ERV-K insert (subgroup LTR5Hs) participates in gene activation in primordial germ cells This ERV-K insert is on chromosome 1, near to the FHAD1 gene. Identified by Xiang et al. (2022), ref. 12.
Transposable Elements
Hundreds of types and subtypes of TE have colonised primate genomes. A class of TE known as SVA elements has arisen only in great apes, and features of their genetic sequence predispose them to participation in gene-regulating functions. Evidence suggests that a subset of SVA elements influence the function of nerve cells. One such element is located between two genes that specify the TRPV1 and TRPV3 proteins. These are ion channels responsive to heat and (in the case of TRPV1) to capsaicin, the pain-eliciting component of chilli peppers, and are also implicated in inflammatory responses. This SVA element appears to regulate the expression of TRPV3 (Figure 3). If the SVA element is deleted experimentally, the activity of the TRPV3 gene is reduced.[14]
The insertion site of this SVA element is shown in Figure 3. It was spliced into the DNA of a human ancestor after the human lineage separated from the chimp/bonobo lineage. The undisturbed target site is present in all other primate species and in the colugo (or flying lemur, a non-primate) and is perfectly preserved in all other apes. This is a human-specific SVA insert, arising in a discrete event in human history, and it will exert human-specific effects.
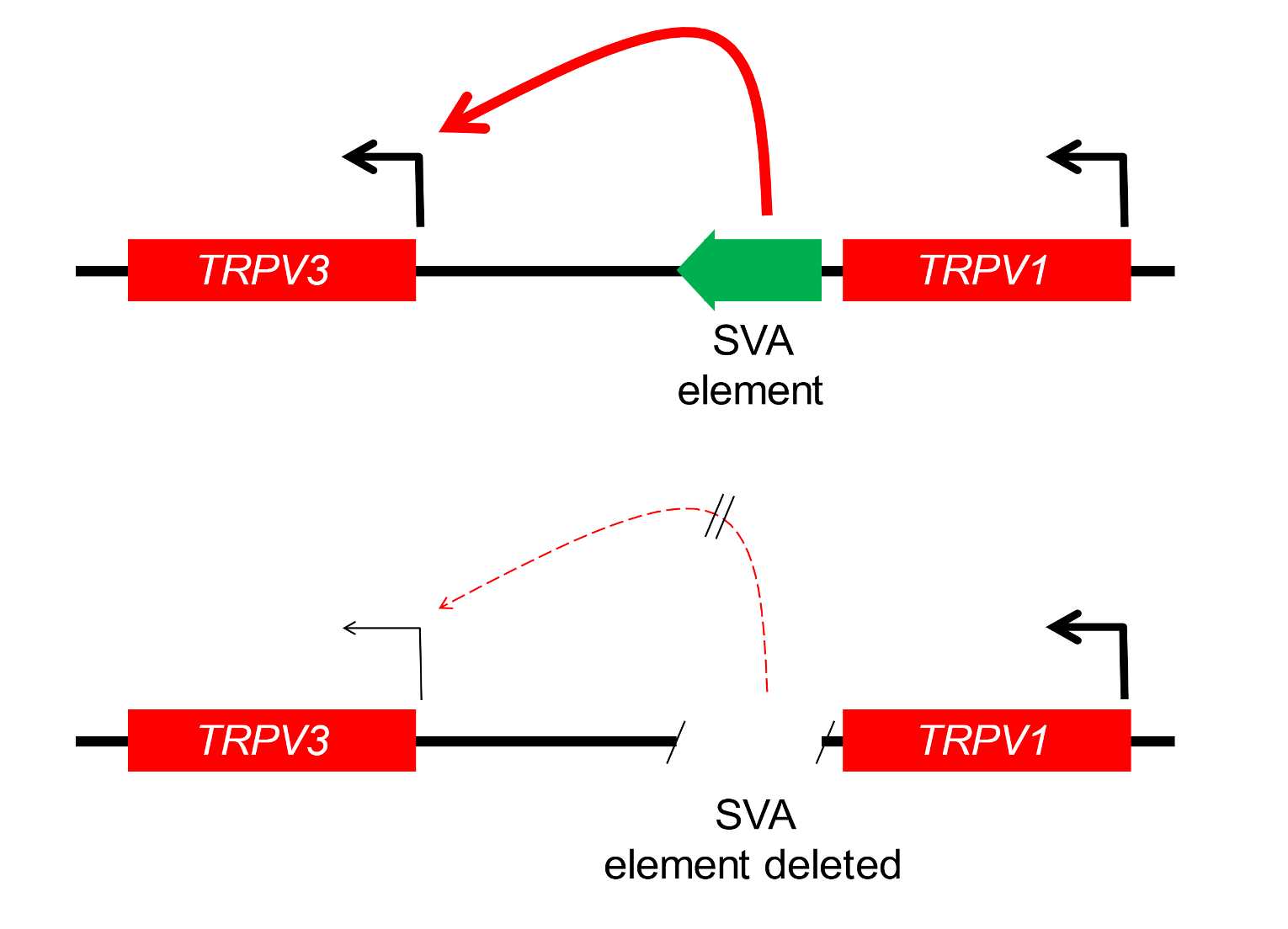
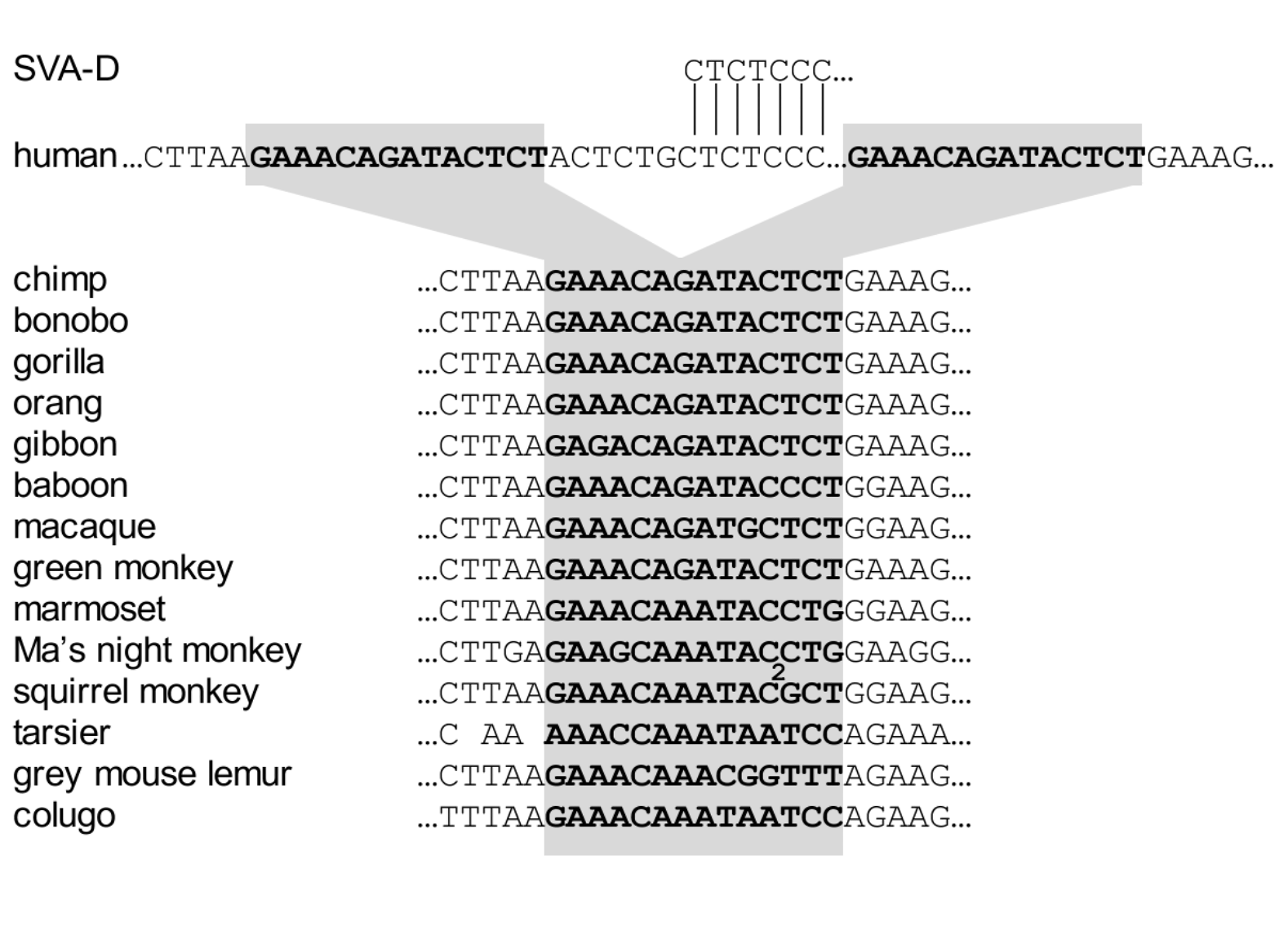
Figure 3. An SVA-D insert that regulates the TRPV3 gene involved in neural function Above: The SVA element (green arrow) activates the TRPV3 gene. If the SVA element is deleted by an experimental procedure (genome editing), TRPV3 gene activity is suppressed. Below: The SVA insertion event occurred since our last common ancestor with chimps. SVA-D sequence is from the Dfam database. This insert was identified in Price et al. (2021), ref. 14.
SVA elements may also drive gene expression in pluripotent stem cells (along with the ERV inserts described above).[15] When pluripotent stem cells become specialised as progenitor cells of the hippocampus (a brain region implicated in learning and memory), ERVs and SVA elements feature as open sites effecting gene regulation. In particular, human-specific SVA elements are associated with changes in gene expression in human relative to chimp cells.[16]
Apes are anomalous relative to other primates for their extended lifespan and increased body size. These features are correlated. A small cohort of SVA elements has been implicated in the regulation of genes that contribute to extended lifespan.[17]
Another characteristic of apes is that they lack a tail. During the development of most mammals, the formation of a tail is initiated by a gene-regulating protein known as brachyury (for which the gene symbol is TBXT). In various mammals, TBXT mutations are known to lead to abnormal tail structure.
In apes, two instances of a TE class known as Alu elements occur in the TBXT gene and may have played an initiating role in tail loss. One of these Alu elements has been present since a common ancestor of all the simian primates (monkeys and apes). It is located (blue arrow, Figure 4) between the fifth and sixth segments of the TBXT gene.
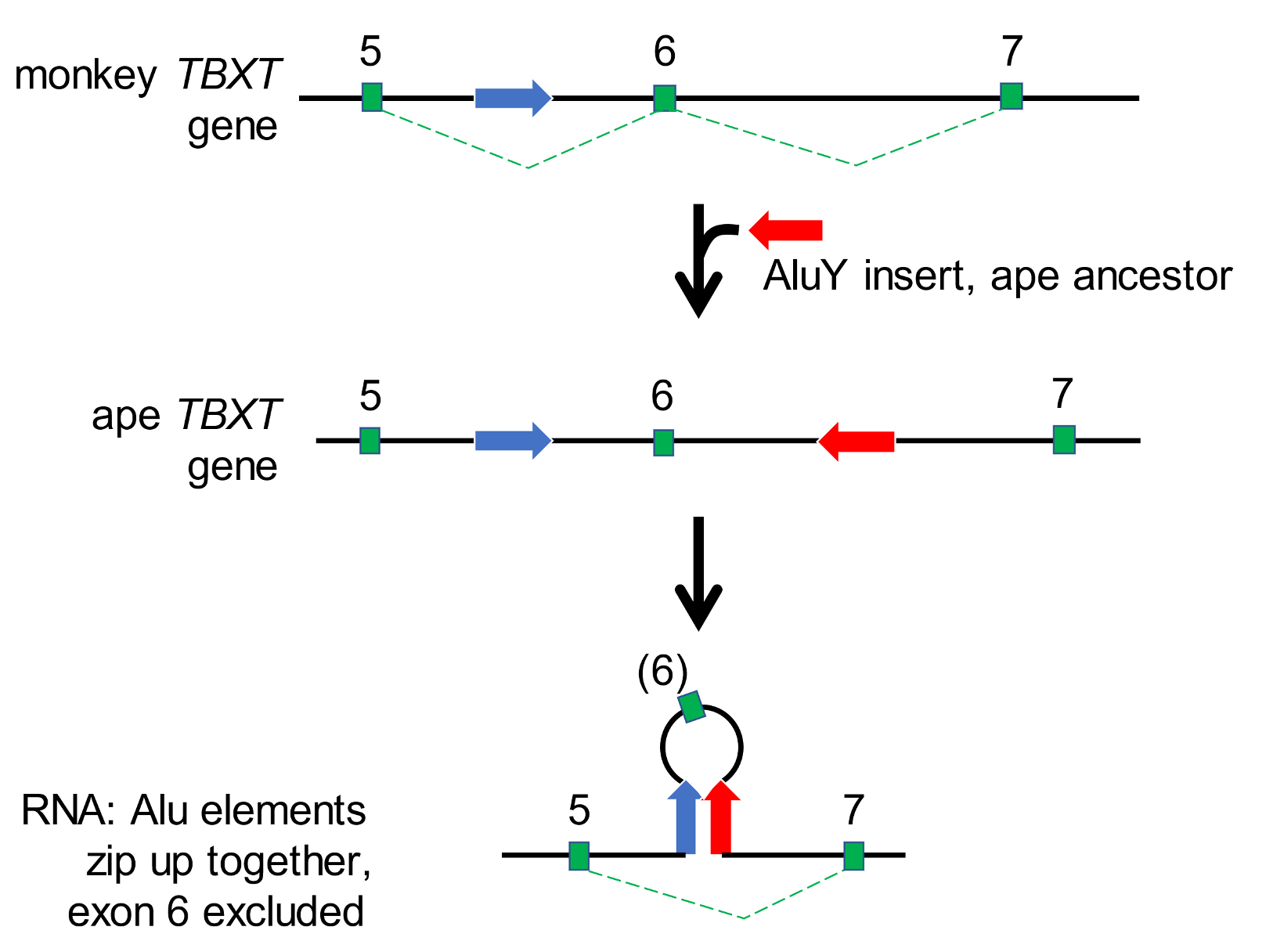
Figure 4. Alu elements suppress activity of the TBXT gene needed for tail development The figure shows the arrangement in monkey DNA (with an Alu element between TBXT gene segments 5 and 6), in ape DNA (with a second Alu element appearing between segments 6 and 7), and in ape RNA, in which the blue and red Alu elements zip up together (hybridise), excluding the sixth segment of the TBXT gene in a loop. The protein made from such RNA molecules will lack amino acids encoded by this segment and will be defective. Early events in tail formation will be suppressed. From Xia et al. (2021), ref. 18.
A second Alu element (red arrow, Figure 4) was added to the primate genome between TBXT segments six and seven in an ancestor of all the apes (including humans). The insertion site of the more recent Alu element is indicated in the alignments of the ape genome sequences (Figure 5). Monkeys, prosimians (tarsier, galago, aye-aye, lemur), and even some non-primates (whales and deer) retain the undisturbed target site.
Once the two Alu elements were in place, they could zip together in an RNA molecule, and sequester TBXT segment six in a loop structure (Figure 4). In this situation, the TBXT protein would be made without the structural component encoded by segment six, and it would possess aberrant activity. In mice, the experimental removal of segment six from the TBXT gene led to abnormal tail formation in many cases. It has been proposed that the Alu-mediated TBXT abnormality comprised a first step in tail loss, and that subsequent genetic events ensured permanent taillessness.[18]
This seems to be a case of evolution by reduction—a case of less is more. An ape-specific Alu element destabilised the TBXT gene and inhibited its activity, so that the tail failed to develop. Loss of the tail may have promoted or expedited the development of bipedality and liberated hominoid forelimbs to engage in delicate manipulations such as are required in the use of tools, fabrication of artefacts, and writing.

Figure 5. An Alu insert that may interfere with a gene TBXT needed for tail development The Alu-Y element is found in all apes. The undisturbed target site is present in monkeys and prosimians, and also survives in Antarctic Minke whale, orca, red deer (sequences from which are not shown).
Alu elements are well-known to influence the readout of genes into which they insert. Hundreds of cases of Alu-modified genes active in the frontal cortex of human brain have been documented. The implications of these are currently unknown.[19] Overall, these discoveries show that randomness in genetic operations—mutational events—can create new information and underlie evolutionary changes that have led to characteristic phenotypes of the human species. Such mutations must occur in the context of constraining selection.
The genomes of essentially all organisms seem to entertain diverse populations of mutagenic ERVs and TEs. The ubiquity of such genetic parasites may seem counter-intuitive, given their propensity to cause disease. But over evolutionary timescales, ERVs and TEs may generate the genomic flexibility that is required to enable evolution to proceed. Indeed, exposing organisms to stressful conditions may increase the activity of such agents and promote evolvability.[20] Levels of randomness may be tuneable. The generation of randomness (in the context of rational selection) is a profoundly efficient way to navigate through environmental challenges.
The necessity of randomness for our daily survival is shown by our adaptive immune system. The production of antibodies requires that antibody genes undergo elevated levels of random mutagenesis (in this case, small changes in DNA sequence), followed by selection of those variants that confer the greatest advantage for our survival—and in a timeframe of weeks. In each developing B cell clone, mutations in antibody genes generate a range of antibody proteins. The presence of antigen selects for those cells that produce antibodies with the tightest fit for the inducing antigen. Immunity thus demonstrates the power of variant generation (mutation) with natural selection. The evolution of a single clone of antibody-producing cells following COVID19 immunisation is depicted in Figure 6.[21]
The development of Darwinian mechanisms of antibody generation also indicates that random mutagenesis in the context of lawful selection can be deeply purposive. Mechanisms of evolution need not be dressed up in an ateleological (purpose-denying) metaphysical garb.[22]
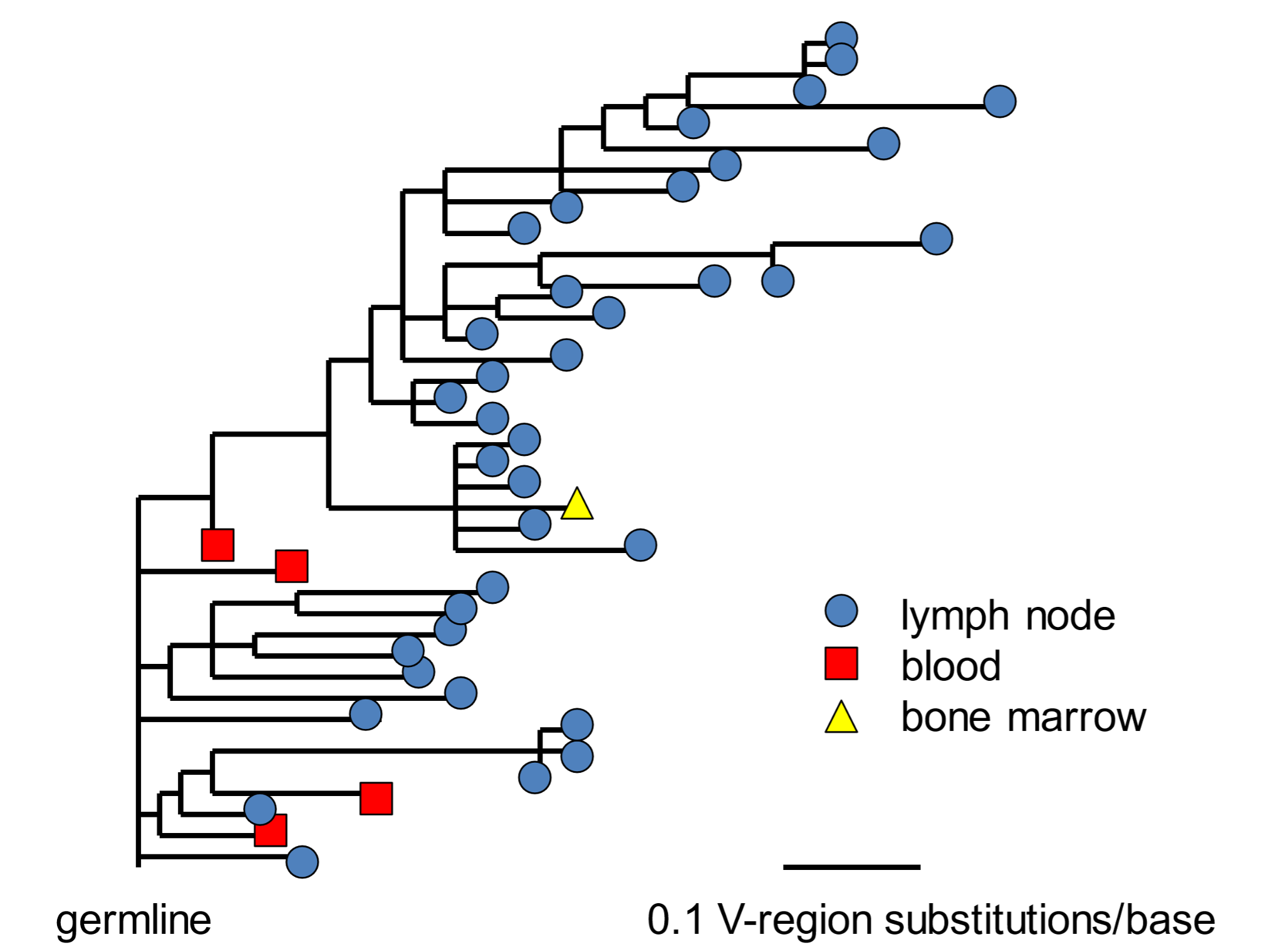
Figure 6. Evolution of a clone of B cells making anti-SARS-CoV-2 antibodies An evolutionary tree of a single clone of antibody-forming cells between four and 29 weeks following immunisation with SARS-CoV-2 vaccine. Genes encoding antibodies are subject to random mutagenesis followed by selection of those that best fit the viral (spike) antigen. The length of the horizontal lines indicates the number of DNA base change mutations (substitutions) as indicated by the scale bar. The V (or variable) region of an antibody molecule is about 110 amino acids long, undergoes high rates of mutation, and provides the interface that binds to target antigens. Abstracted and adapted from Kim, Zhou, Horvath et al. (2022), ref. 21.
Processes that are random at the micro-level emerge into highly ordered and predictable phenomena at the macro-level. Physicist Tom McLeish has described how randomness or chaos of molecular processes (like Brownian motion) give rise, in the context of lawful constraints, to emergent order in living cells. The science of statistical mechanics provides an understanding “of how predictable, ordered structure and behaviour at the macroscopic scale emerged from a microscopic world of disorder.” McLeish considers that this insight “is one of the most remarkable achievements of physics over the last century and a half.”[23]
Christians may gladly accept that “local chaos can give rise to large-scale structure when there are additional constraints, that creation harnesses the power of random forces without suppressing them, but rather by directing them into paths and processes, even extending them to the processes of life itself.” Random mutations can give rise to the macroscopic properties of the developed organism.[24] ERVs and TEs in all their stochasticity have contributed to a creature that is human rather than, say, chimp. The evolution of life itself can be seen to fall “into the category of ordered large-scale structure emergent from random small-scale dynamics.”[25] Random mutations can underlie predictable trajectories of evolution.
McLeish has extended the theme of “chaos to emergent order” to the whole of life. The biblical character of Job questioned the random events that afflicted him at the micro-level of his own existence. God’s answer pointed him to the emergent order and beauty manifested in the universe at the macro-level. There seems to be an apparent lack of control in the “microscopics” of mutation and other disruptive influences, but we (with Job) should recognise how such creative energies “unfold the possibilities of the created order.”[26]
Creation
The representative studies discussed above indicate that particular genetic events, describable at atomic resolution, have contributed to aspects of our humanness. Random events (in the appropriate context) can indeed generate new information and, during human development, modify regulatory circuitry. A long history of such events has led to the advent of Homo sapiens.
Such discoveries provide some clarity to our earlier question pertaining to a creator. Evolutionary process is complete in its own mechanistic terms. A god conceived as a component of DNA biochemistry is indeed redundant. Matter has within itself the potential to complexify. But the question of a creator has more to do with why there should be matter, why it should possess potentiality, and why there should be a drive to complexification. As cosmologist Heino Falcke has stated, scientists “have come better to understand the rules of the game in the universe, but where the game and where the rules come from, this we haven’t answered.”[27]
The elucidation of biochemical mechanisms underlying evolution has nothing to say about God as creator. Such a God can be considered only as the source of the entire system. If we are to think biblically, we must recognise that the atoms constituting DNA, the characteristically random but intelligible behaviour[28] intrinsic to mutagenic agents (such as ERVs and TEs), and the context in which mutations undergo selection, are all components of created reality.
Biblical creation is expressed by the Hebrew word bara (which is used exclusively of God’s action) and by many broadly synonymous terms.[29] Creation/bara and its synonyms essentially describe the authority of God over creation, and indicate that physical entities and the processes in which they engage are conceived, willed, and effected by God.[30] Such terms encompass God’s authority over familiar phenomena that are regarded as wholly natural (such as the wind and rain).[31] Creation/bara denotes divine sovereign effectuation,[32] a divine bringing into being, and relates to happenings (judgment and redemption), conditions (light and darkness), acts of God’s saving righteousness, and the transformation of a person’s heart to a state compatible with God’s holiness.[33]
One of these meanings has to do with existence, the gift of being.[34] The biblical concept of creation implies the traditional idea of creatio ex nihilo—creation out of nothing.[35] The famous “Let there be” statements of the first Genesis creation story[36] have everything to do with the conferral of being. That God’s creative work is to give being is reflected in one of the great creation Psalms: “For he spoke, and it came to be; he commanded, and it stood firm.”[37]
In Athens, St Paul quoted a pagan philosopher, Epimenides, with approval: For in God “we live and move and have our being.”[38] Paul wrote in his magnum opus, the letter to the Roman Christians, that God’s “command brings into being what did not exist.”[39] And Paul emphasised the all-encompassing scope of God’s work in an outburst of praise, “For all things were created by him, and all things exist through him and for him. To God be the glory forever! Amen.”[40] In the heavenly vision of St John,[41] God is worshiped for the gift of being, of existence: “You are worthy, our Lord and God, to receive glory and honour and power, for you created all things, and by your will they were created and have their being.”
The gift of existence may be variously nuanced. Walton informs us that Western thought tends to understand creation, being, or existence in physical terms.[42] In contrast, the ancient Hebrews understood that something existed “by virtue of its having a function in an ordered system”—in particular, according to how it related to society and culture.[43] People’s ontology, their understanding of reality, focused on what they believed to be its most significant feature.[44]
If we apply Walton’s proposal to genetics, we might suggest that divine creation pertains not only to the existence of biomolecules such as DNA, but to the function capacities of DNA. It is an extraordinarily stable repository of information (continually responsive to, and updated by, environmental influences that impinge upon organisms), with sufficient mutability to be a vehicle for the development of spectacularly diverse life forms, and the genetic substrate of at least one creature that could respond in adoration to God’s address.
Confusing Creation
Some materialist authors claim that biological evolution justifies atheistic belief. This is absurd, for how can we imagine a history—any history—as being an alternative to the conception that the cosmos in which it occurs is created, ordered, and sustained by God? There can be absolutely no incongruity in accepting the findings of historical science (including evolutionary genetics) and believing that everything accessible to science is ordained by God.
Some materialistically minded science writers have proposed that, if the cosmos was proven to emerge from a prior state (say the quantum vacuum), then the need for a creator is thereby obviated. For example, the cosmologist Lawrence Krauss has promoted the idea that the universe arose from “almost nothing”—where the pre-existing “almost” includes the laws of physics, the spectrum of fundamental particles,[45] and the provision of highly structured quantum fields. “In a Christian understanding, that provision would be the continuing act of the Creator.”[46]
David Bentley Hart chides such materialists for their crude verbal trickery. The transition from any preexisting physical reality to our own familiar universe is purely a change from one state to another and has no relevance to the question of being.
Hart has said that “all physical events … are embraced within the history of nature, which is to say the history of what already has existence. The question of existence, however, concerns the very possibility of such a history.” In other words, the biblical concept of creation encompasses the whole of physical reality. “Any quantum fluctuation [within an existing quantum system] that produces, say a universe is a new state within that system, but not a sudden emergence of reality from nonbeing.”[47]
Stephen Hawking proposed that the cosmos is like a fuzzy spacetime egg without a beginning or an end. He asked whether such a boundary-less universe allowed any room for a creator. John Polkinghorne responded that Hawking’s proposal was scientifically interesting but theologically inconsequential, for God is present in every place, “as the sustainer of the self-contained spacetime egg and the ordainer of its quantum laws.” God is not limited by boundaries.[48] Hart quotes the theologian E. L. Mascall with approval: God is not “just one item, albeit the supreme one, in a class of beings,” but is rather “the source from which their being is derived.”[49]
We dare not confuse biblical creation with any physical or biological process belonging to the category of evolution. A biblical concept of creation entails that all of reality—every atom, every photon, and every instant of time—is given existence by God. As Douglas Spanner said in 1987, “in the Bible, the creative aspect of God’s activity … is never linked to a particular time, place, process or material; the act is seen rather as an unanalyzable movement out of the infinity of God’s thoughts into the finiteness of time and space and all that fills them.”[50] Evolution then (whether it describes the development of the cosmos, of life, or of the piano) is a created process. Creation and evolution cannot be alternatives. Evolutionary history is a process situated within God’s created world.
The astronomer Howard Van Till has provided useful distinctions between the scientific investigation of the world and the biblical idea of creation. Science focuses on the cosmos in terms of its coherent properties, its lawful behaviour, and its authentic history. The Judeo-Christian idea of creation, however, considers the cosmos as an ever-dependent reality in relation to its creator. God is its Originator, Preserver, Governor, and Provider.[51] Thus, while science investigates the relationships between component parts of the world, theology in its metaphors describing creation depicts the relationship between the world and God.[52]
Brueggemann has stated that, to Israel, creation was “covenantally ordered; that is, formed for continuing interaction of gift and gratitude, of governance and obedience.” God’s action in creation is never the expression “of raw, sovereign power,” but is rather characterised by “covenantal, ethical intentionality.”[53] Discourse on evolution is stringently limited to physical phenomena, whereas that pertaining to creation expands the vistas to purpose, faithfulness, and hope.
God’s Gift of Being: Implications
A criterion of the validity of a scientific hypothesis is that it should be fruitful—that it should throw unexpected light on other questions relating to physical reality. A valid theological insight should have the same capacity to make sense of diverse questions of our experienced reality—albeit issues that are personal (not mechanistic) ones. Of relevance to the current discussion, the biblical concept of creation enlightens and enriches central aspects of our humanity.
First, the concept of creation has provided the worldview conducive to the development of science.[54] Three senior physicists have written: “[Christianity] practically invented science. Or at least, the striking progress of science in the modern era had many of its roots in Christian theistic belief, and for four hundred years the Christian community has largely nurtured science and done it well.”[55] To qualify this statement, it should be stressed that humanity at large has engaged in careful observation of the natural world, but the biblical depiction of the divine nature (for example, God’s authority, wisdom, faithfulness, freedom, goodness, and glory) has provided presuppositions that enabled science to flourish. The understanding of our world as creation has been hugely fruitful for the growth of science and the benefits flowing from it. That the biblical concept of creation has facilitated the development of science is evidence that the Hebraic understanding of the creator God entails a singularly valid purchase on reality. There is something special about biblical ontology.
Second, our status as created beings gives us identity and dignity. A scientist’s perusal of human DNA sees that of just another ape. The human DNA sequence is most similar to that of chimps, followed by those of gorillas and orangutans (Figures 1, 2, 5). Genes in these related species are similarly interspersed amongst a jumbled concatenation of ERVs and TEs (over 99% of which are shared between human and chimp genomes). But we are not an inconsequential byproduct of selfish DNA. We are hominoid primates valued, loved, and called by God. Our physicists write that “we are not just forced into being but called into being. That is, we—all humans—and the other animals too, to a more limited extent, are not just forced into existence by the inexorable and blind processes of the physical world … We are called, as people, by one who so calls. We are talked into talking, loved into loving, and forgiven into forgiving.”[56]
Materialistic writers loudly proclaim their creed that the universe is devoid of ultimate significance or purpose. They deny that goodness, compassion, or justice are written into the structure of reality. Such a nihilistic creed is fine for wealthy celebrities. But one would expect it to be inimical to the wellbeing of people struggling to find their identity in the fickle currents of contemporary ideologies. In principle, the understanding of genetic process cannot speak to the mystery of human uniqueness as Homo credens—believing human. To our physics professors, “the analysis and description of a process cannot, logically, even address the issue of the overall meaning and purpose of that process, nor can it address what made it possible for that process to happen in the first place.”[57] The dissection of genetic events in our evolution is deeply fascinating at an intellectual level, but it is the knowledge of God as the source and goal of our being that guarantees our inalienable value as persons. Cosmologist Heino Falcke has said that science tells us how small we are; theology tells us how valuable we are.[58]
We were formed in utero by genetic programmes (in dependence on environmental conditions) that were constructed during evolutionary history at least partially by the stochastic activities of retroviruses and transposable elements. But these impersonal processes have enabled us to enter into the dimension of the personal and the relational.
Natural selection, as it were, discovered [the very concept of personhood]; it does not cause it. Natural selection favoured the eventual emergence of complex creatures able to embody personhood; when this emerged, it could not do other than embody what personhood is. The “mystery of our existence” is, in fact, very much about the nature of personhood. The nature of personhood is not explained by the physical process through which it became embodied in the physical world.[59]
Third, the idea of creation provides reassurance in the face of pervasive chaos. To Stanley Jaki, the faith of ancient Israel “emphasised the idea of the utter dependence of everything on one single Being.” In the second creation story of Genesis 2, “there is only one effective cause”—God—who “is not challenged or complemented” by anyone or anything else.[60] The Hebrews had a highly confident vision of nature, as a home for humanity, where humans could develop their unique potentialities. The cosmos was “not an agglomerate of capricious events and processes” subject to the domination of unpredictable and dark forces.[61] Jaki had in mind the connection between Israel’s faith in the creator God and the later development of a scientific vision of nature. But Israel’s faith in the covenantal God also underlays attitudes of virtue and positivity in which human wellbeing could flourish.
To Israel, creation was made for glad dependence on God and fruitful obedience to God.[62] Creation faith focuses our attention on personal realities rather than mechanistic speculation or explanation. “It invites wonder, awe and gratitude that life—Israel’s life, human life—is situated in the midst of a reliable generosity that precedes all human effort.”[63] Creation addresses human wellbeing and flourishing in a way that scientific categories in principle never can. As the wondering Hebrew poet expressed it:
When I consider the heavens, the work of your fingers,
the moon and the stars which you have set in their place,
what are mere mortals that you are mindful of them,
human beings that you care for them?[64]
Fourth, the rich and fertile Hebraic concept of creation gives credence to the hope of future development, of perfectibility, in God’s reality. The splendour of this world has been attained at a concomitant cost. Biological history has issued in disease, suffering, and death. Human history is a story of barely mitigated savagery. The belief that creation comes from a good God generates the haunting hope that creation is not complete[65] and that a cosmos freed from its slavery to decay may be anticipated. A different sort of reality, of cosmos, must be expected in which suffering and human savagery can no longer exist.
Brueggemann (citing Jon Levinson) has said that “something untamed and destructive” remains loose in the world; and that it still needs to be brought under the rule of God. “Creation faith is the summons and invitation to trust” in this God, “even in the face of day-to-day, palpable incursions of chaos.” The testimony of Israel is that God “can be trusted in the midst of any chaos, even that of exile and finally that of death.”[66]
Creation faith precedes and enables the anticipation of a new creation, that this world will be transformed into one in which (through God’s own sacrificial involvement in Jesus) the suffering and evil endemic to current reality will be extirpated.[67] “God was in Christ reconciling the world [κόσμον] to himself.”[68] As an aspect of this, human savagery will be replaced by the creation of a new humanity.[69] And individuals will be (are being) created anew as new people.[70] For “God has made us what we are, and in our union with Christ Jesus he has created us for a life of good deeds, which he has already prepared for us to do.”[71] Of course, a proposal or idea is not true simply because it is convenient. It is true because it makes sense of the real world we experience.
In conclusion, we can confidently accept both the molecular genetic evidence of our development through evolutionary history and the fruitful biblical assertion that we are created beings. The scientific and theological perspectives on our nature are complementary and highly enriching. Together they provide a coherent (although still incomplete) understanding of our nature as evolved hominoid primates who find their fulfilment in encountering God as creator and redeemer.
The author reports there are no competing interests to declare.
Received: 06/12/22 Accepted: 02/03/23 Published: 07/03/23
[1] For other fascinating examples of novel features, see my Human Evolution: Genes, Genealogies and Phylogenies (Cambridge: Cambridge University Press, 2013) and Evolution and Eschatology (Eugene, OR: Wipf and Stock, 2021). Retroviruses and transposable element insertions are only one class of mutation that has formed our genome. But I have focused on these agents because each event is essentially unique, the prior state and the derived mutated state in the genome are known, the enzyme-catalysed mechanisms by which they arise have been elucidated, the functional consequences of these mutagens are amenable to investigation, and (to my mind) they generate fascinating stories![2] See Graeme Finlay, “Evolution as History: Phylogenetics of Genomes and Manuscripts,” Christian Perspectives on Science and Technology, New Series 1 (2022): 150–174.
[3] For example, the role of SVA elements in genetic disease is discussed in Abigail L. Pfaff, Lewis M. Singleton, and Sulev Kõks, “Mechanisms of Disease-Associated SINE-VNTR-Alus,” Experimental Biology and Medicine 247 (2022): 756–764.
[4] For reviews, see R. Michael Roberts, Toshihiko Ezashi, Laura C. Schultz et al., “Syncytins Expressed in Human Placental Trophoblast,” Placenta 113 (2021): 8–14; Kazuhiko Imakawa, Kazuya Kusama, Tomoko Kaneko-Ishino et al., “Endogenous Retroviruses and Placental Evolution, Development, and Diversity,” Cells 11 (2022): 2458.
[5] The gene encoding suppressyn resides on chromosome 21, which in Down’s syndrome is present in an extra copy. Excessive production of suppressyn because of trisomy 21 may cause placental abnormalities in Down’s pregnancies by inhibiting cell-cell fusion and syncytiotrophoblast formation. See Jun Sugimoto, Danny J. Schust, Tomomi Yamazaki, and Yoshiki Kudo, “Involvement of the HERV-Derived Cell-Fusion Inhibitor, Suppressyn, in the Fusion Defects Characteristic of the Trisomy 21 Placenta,” Scientific Reports 12 (2022): 10552.
[6] John A. Frank, Manvendra Singh, Harrison B. Cullen et al., “Evolution and Antiviral Activity of a Human Protein of Retroviral Origin,” Science 378 (2022): 422–428.
[7] Arnon Plianchaisuk, Kazuya Kusama, Kiyoko Kato et al., “Origination of LTR Retroelement-Derived NYNRIN Coincides with Therian Placental Emergence,” Molecular Biology and Evolution 39 (2022): msac176.
[8] For a review, see Anna D. Senft and Todd S. Macfarlan, “Transposable Elements Shape the Evolution of Mammalian Development,” Nature Reviews Genetics 22 (2021): 691–711.
[9] Sanna Vuoristo, Shruti Bhagat, Christel Hyden-Granskog et al., “DUX4 is a Multifunctional Factor Priming Human Embryonic Genome Activation,” iScience 25 (2022): 104137; Kosuke Hashimoto, Eeva-Mari Jouhilahti, Virpi Tohonen et al., “Embryonic LTR Retrotransposons Supply Promoter Modules to Somatic Tissues,” Genome Research 31 (2021): 1983–1993.
[10] These are primitive cells found in the early embryo that have an unlimited ability to proliferate (given the right conditions) and the potential to produce all the specialised cell types of the mature organism.
[11] Thomas A. Carter, Manvendra Singh, Gabrijela Dumbović et al., “Mosaic Cis-Regulatory Evolution Drives Transcriptional Partitioning of HERVH Endogenous Retrovirus in the Human Embryo,” eLife 11 (2022): e76257.
[12] Xinyu Xiang, Yu Tao, Jonathan DiRusso et al., “Human Reproduction is Regulated by Retrotransposons Derived from Ancient Hominidae-Specific Viral Infections,” Nature Communications 13 (2022): 463; Jumpei Ito, Yasunari Seita, Shohei Kojima et al., “A Hominoid-Specific Endogenous Retrovirus may have Rewired the Gene Regulatory Network Shared between Primordial Germ cells and Naïve Pluripotent Cells,” PLoS Genetics 18 (2022): e1009846.
[13] ERVs and TEs continue to bind gene-regulatory proteins through later stages of development. LTR5Hs binds pluripotency-sustaining proteins (such as KLF4) in stem cells of early embryos, and cell lineage-specific regulatory proteins (such as SOX17, GATA6 and TBXT) in subsequent stages. See Julian Pontis, Cyril Pulver, Christopher J. Playfoot et al., “Primate-Specific Transposable Elements Shape Transcriptional Networks During Human Development,” Nature Communications 13 (2022): 7178. In the maternal part of the placenta (the uterine decidua), many classes of ERV and TE provide binding sites for gene-regulatory proteins more frequently than expected by chance, including 62.8% of the binding sites for the progesterone (pregnancy hormone) receptor. Many sites transform from gene-repressive to -enhancing depending on epigenetic controls. See Katelyn Mika and Vincent J. Lynch, “Transposable Elements Continuously Remodel the Regulatory Landscape, Transcriptome, and Function of Decidual Stromal Cells,” Genome Biology and Evolution 14 (2022): evac164. At least one endogenous retrovirus of the ERV-K subclass is genetically active in each of fifty-four adult tissues as well. Whether such ERV-K activity is random noise or implies hitherto unsuspected functionality is not known. Aidan Burn, Farrah Roy, Michael Freeman, and John M. Coffin, “Widespread Expression of the Ancient HERV-K (HML-2) Provirus Group in Normal Human Tissues,” PLoS Biology 20 (2022): e3001826.
[14] Emma Price, Olympia Gianfrancesco, Patrick Harrison et al., “CRISPR Deletion of a SVA Retrotransposon Demonstrates Function as a cis-Regulatory Element at the TRPV1/TRPV3 Intergenic Region,” International Journal of Molecular Sciences 22 (2021): 1911.
[15] Samantha M. Barnada, Andrew Isopi, Daniela Tejada-Martinez et al., “Genomic Features Underlie the Co-option of SVA Transposons as Cis-Regulatory Elements in Human Pluripotent Stem Cells,” PLoS Genetics 18 (2022): e1010225.
[16] Sruti Patoori, Samantha M. Barnada, Christopher Large et al., “Young Transposable Elements Rewired Gene Regulatory Networks in Human and Chimpanzee Hippocampal Intermediate Progenitors,” Development 149 (2022): 200413.
[17] Daniela Tejada-Martinez, Roberto A. Avelar, Inês Lopes et al., “Positive Selection and Enhancer Evolution Shaped Lifespan and Body Mass in Great Apes,” Molecular Biology and Evolution 39 (2022): msab369.
[18] Bo Xia, Weimin Zhang, Aleksandra Wadzinska et al., “The Genetic Basis of Tail-Loss Evolution in Humans and Apes,” BioRxiv https://doi.org/10.1101/2021.09.14.460388.
[19] Liliana Florea, Lindsay Payer, Corina Antonescu et al., “Detection of Alu Exonization Events in Human Frontal Cortex From RNA-Seq Data,” Frontiers in Molecular Biosciences 8 (2021): 727537.
[20] Elizabeth A. Mojica and Dietmar Kültz, “Physiological Mechanisms of Stress-Induced Evolution,” Journal of Experimental Biology 225 (2022): 243264.
[21] Wooseob Kim, Julian Q. Zhou, Stephen C. Horvath et al., “Germinal Centre-Driven Maturation of B Cell Response to mRNA vaccination,” Nature 604 (2022): 141–145.
[22] Graeme Finlay, “The Immune System: Unity in Community,” Science and Christian Belief 34 (2022): 29–49.
[23] Tom McLeish, “Evolution as an Unwrapping of the Gift of Freedom,” Scientia et Fides 8 (2020): 43–64; quote from p. 48.
[24] McLeish, “Evolution,” 48–49.
[25] McLeish, “Evolution,” 49.
[26] McLeish, “Evolution,” 58.
[27] Heino Falcke, Light in the Darkness: Black Holes, the Universe and Us (London: Wildfire, 2021), 285.
[28] As noted above, randomness is ordered, as described by statistical mechanics. See Tom McLeish, Faith and Wisdom in Science (Oxford: Oxford University Press, 2014), 100–101.
[29] Howard J. Van Till, Robert E. Snow, John H. Stek, and Davis A. Young, Portraits of Creation (Grand Rapids, MI: Eerdmans, 1990), 208–211.
[30] Van Till et al., Portraits, 213.
[31] Van Till et al., Portraits, 214, 216.
[32] Van Till et al., Portraits, 218–219, 221.
[33] Van Till et al., Portraits, 208.
[34] Van Till et al., Portraits, 213.
[35] Adrio Konig, New and Greater Things (Pretoria: UNISA, 1988), 102–104, 120 (allowing that a diversity of metaphors describing creation is used in Scripture).
[36] Gen 1:3, 6, 14.
[37] Ps 33:9, NIV.
[38] Acts 17:28, NIV; or exist, GNT.
[39] Rom 4:17.
[40] Rom 11:36, GNT.
[41] Rev 4:11, NIV.
[42] John H. Walton, The Lost World of Genesis One (Downers Grove, IL: IVP, 2009), 23–25.
[43] Walton, Lost World, 26, 35, 53; and associated Chapters 4–6.
[44] Walton, Lost World, 28.
[45] Roland Ashby, Chris Mulherin, John Pilbrow, and Stephen Ames, A Reckless God? (Reservoir, Victoria: Morning Star, 2018), 44–45; comment on fundamental particles is from Professor Jeff Tallon, personal communication.
[46] John Polkinghorne, Science and Creation (London: SPCK, 1988), 60.
[47] David Bentley Hart, God: Being, Consciousness, Bliss (New Haven: Yale University Press, 2013), 98.
[48] John Polkinghorne, Science and Christian Belief (London: SPCK, 1994), 73.
[49] Hart, God, 108.
[50] Douglas C. Spanner, Biblical Creation and the Theory of Evolution (Exeter: Paternoster, 1987), 35.
[51] Howard Van Till, The Fourth Day (Grand Rapids: Eerdmans, 1986), xiii–ix; 62–65.
[52] Van Till, Fourth Day, 64.
[53] Walter Brueggemann, Theology of the Old Testament (Minneapolis, MN: Fortress, 1997), 157–58.
[54] Christopher Kaiser, Creation and the History of Science (London: Marshall Pickering, 1991); Harold Turner, The Roots of Science (Auckland: DeepSight Trust, 1998); Mark Worthing, Unlikely Allies: Monotheism and the Rise of Science (Eugene, OR: Wipf and Stock, 2019); Graeme Finlay, God’s Gift of Science: Theological Presuppositions Underlying Exploration of the Natural World (Eugene, OR: Wipf and Stock, 2022).
[55] Andrew Briggs, Hans Halvorson, and Andrew Steane, It Keeps Me Seeking (Oxford: Oxford University Press, 2018), 9.
[56] Briggs et al., It Keeps Me Seeking, 2.
[57] Briggs et al., It Keeps Me Seeking, 186.
[58] Falcke, Light in the Darkness, 290.
[59] Briggs et al., It Keeps Me Seeing, 188.
[60] Stanley Jaki, Science and Creation (Edinburgh: Scottish Academic Press, 1986), 139–140.
[61] Jaki, Science and Creation, 148, 150.
[62] Brueggemann, Theology, 149.
[63] Brueggemann, Theology, 156.
[64] Ps 8:3–4.
[65] Konig, New and Greater Things, 159.
[66] Brueggemann, Theology, 159.
[67] Isa 65:17; 2 Pet 3:13; Rev 21:1.
[68] 2 Cor 5:19.
[69] Eph 2:15; Heb 8:8–13.
[70] 2 Cor 5:17.
[71] Eph 2:10.