
Christian Perspectives on Science and Technology, New Series, Vol. 3 (2024), 87–135
Abstract: Evolutionary biology is regarded with suspicion in some theological circles. However, the era of DNA sequencing, allowing linear side-by-side comparisons of our genome with those of related species, has revealed how our genetic “text”—that is, our genome—has incrementally developed, and has provided compelling insights into our evolutionary history. This paper presents examples of mutations in our genetic text that have generated features of our characteristically human biology. It is suggested that our genetic text be thought of as the Primal Testament, analogously to the Old and New Testaments that describe the history of God’s dealings with humankind. There are clear differences between the Primal Testament and those of the scriptures. The Primal Testament describes an impersonal and nonmoral history. But the three testaments have in common their witness to God’s purposes, their accounts of God’s creation of new realities (biological organisms, Israel, and the church), and their depictions of richer conceptions of life (successively, biological, personal/communal, and the Spirit-indwelt zoe aionios). The three testaments all describe contingent histories arising from God’s gift of freedom to the creatures. The histories alike describe ambiguities, suffering (even extinctions), goal-directedness, and incompleteness as they together anticipate God’s consummation of all things in the New Creation.
Biological evolution and divine creation are often discussed as if they were starkly alternative ideas. Indeed, some Christians may express distaste for—or categorical rejection of—the concept of evolution because it is not compatible with the postulate that creation underwent a cataclysmic fall from an originally perfect, idyllic golden age. Moreover, people may assert that evolutionary process is inherently random and wasteful; and decry phylogenetic history as a saga of suffering, death, and mass extinctions. For many, the idea of evolution seems to exclude considerations of an overarching purpose for the cosmos.
But many of the psychologically disturbing features perceived in biological evolution are also present in the histories described in the Old Testament (or Hebrew scriptures) and New Testament. Biblical history is replete with human failure, blind alleys, and arbitrary savagery that is foreign to the nature of God. Evolutionary biology and biblical history share some commonality of character. To John Polkinghorne, there are at least methodological similarities between the historical sciences and biblical studies. The study of the “unique and fragmented records that are the basis for the insights of historico-observational sciences such as evolutionary biology and physical cosmology” comprise the closest analogy amongst the sciences to biblical studies.[1] In the same way as evolutionary biology has been rigorously and painstakingly studied to provide a developing outline of prehuman history, the rational use of the Bible as a source of information for the histories of Israel and of Jesus of Nazareth provides (as Polkinghorne continues) “a foundation for Christian belief that can be defended with intellectual care and scrupulosity.”
The thesis of this paper is that phylogenetic history is written in genomes, and comprises a Primal Testament,[2] albeit in the impersonal and nonmoral language of DNA. Francis Collins refers to the DNA text that constitutes the genome as the language of God.[3] The Primal Testament has its own authority, analogous to that of the biblical testaments written in Hebrew and Greek (Figure 1). The Primal Testament describes how God has ordained and sustained a developing biosphere in the same way as the Old and New Testaments describe the history of God’s deepening engagement with humanity. All three testaments reflect tumultuous histories. They describe the actions of agents created to act freely within the time dimension of the created space-time order.

Figure 1. Authoritative texts describing three formative histories. Languages (left to right) are genetic DNA sequence (from the KRAS gene), Hebrew (Isaiah Scroll, Qumran), and Greek (Codex Sinaiticus). Sources: NCBI (Refseq database) and Wikimedia Commons.
Created history extends far beyond what can be studied in written texts. God is the Author of all histories, even those not documented by one of the detailed texts to which Figure 1 refers. For this reason, an Appendix is added to this paper: my poem entitled “A Freed Man’s Worship” that acknowledges divine authorship of all histories, however they have been reconstructed.
Episodes in Evolutionary History
We consider examples of the types of historical data encountered in evolutionary genomics, as documented in the text written in the language of DNA. They provide details of what happened in the remote past, and provide insights regarding our nature in the experienced present.
Pseudogenes
Our genomes contain thousands of degenerated genes that do not function in the way the corresponding genes do in other species. Such gene relics are called pseudogenes.[4] We will consider two such genes that cannot make their respective proteins. It seems that, at some time in the past, their functions became redundant and in the absence of selective pressure, they started to accumulate inactivating mutations. They appear to be decaying into the genomic background.
Many animals communicate with each other using chemical messengers called pheromones. These regulate mating and other social behaviours. Pheromones activate receptors found on sensory nerve cells in the vomeronasal organ of the nasal cavity. The receptors are of two classes, V1R and V2R. Humans lack a vomeronasal organ, their complements of V1R- and V2R-encoding genes have been drastically reduced and, in all probability, they do not respond to pheromone signals.
A particularly ancient V1R gene (hence, ancV1R) is present in vertebrate genomes from bony fish to humans. However, in humans, other apes, and Old World monkeys, the ancV1R gene has been scrambled by inactivating mutations. In protein-coding segments of DNA, triplets of bases specify which amino acids are added into proteins. In the case of all the great apes, a TTA triplet (specifying the amino acid leucine, L) has mutated to TAA (a stop signal at which the elongation of protein chains is terminated) (Figure 2). The result is a truncated protein that is probably devoid of function. The only way by which the five great ape species could share such a mutation is for it to have arisen in an ancestor from which all are descended. Old World monkeys possess a different mutation (the catastrophic loss of a base), which occurred in an ancestor of these primates.[5]

Figure 2. Inactivating mutations in the ancV1R gene. The gene has been pseudogenised independently in great apes (TTA to TAA, leucine to stop), lesser apes (CAA to TAA, Q to stop, not present in the sequence shown above) and Old World monkeys (GTA triplet reduced to TA, with all subsequent triplets shifted out-of-phase). The single letters above the human and marmoset gene sequences represent the amino acids constituting the encoded protein. Suzuki et al. (2018).
The presence of the ancV1R gene in our genome indicates that we are descended from distant forbears that signalled to each other by pheromones. The interpretation of ancV1R inactivation is that at some time, our ancestors lost the need to communicate through pheromones. If you don’t use it, you lose it. Perhaps the development of an acute sense of vision superseded communication through chemicals.
Insectivores possess enzymes that digest the chitin exoskeletons of insects. It is believed that early mammals were largely insectivorous but with the end of the dinosaur era (66 million years ago), mammals diversified to occupy a multitude of ecological niches. New dietary habits included herbivory and carnivory. Many ceased to depend on insects, and chitin-digesting enzymes (chitinases) were no longer needed for nutrition. Genes for chitinases were not maintained by natural selection and tended to decay. A chitinase gene (CHIA2) relic possesses a stop mutation in all simian primates, including humans (Figure 3). An amino acid-specifying triplet (probably CGA, retained in tarsiers) has mutated into a gene-inactivating stop triplet, TGA. All simians possess the mutant, which arose in a unique event in an ancestor of simian primates.[6]

Figure 3. Inactivating mutation in a chitinase (CHIA2) gene. The gene has been pseudogenised in all simian primates (CGA to TGA, arginine to stop). Emerling et al. (2018).
John the Baptist’s diet of locusts would have provided protein, but he could not have extracted carbohydrates from the chitin that constitutes the locust exoskeleton. The CHIA2 gene in John’s genome had fallen into disrepair millions of generations before his time. His consumption of wild honey provided energy-sustaining sugars.
Transposable Elements
Another major class of genome-disrupting events (mutations) arises from the actions of parasitic units of DNA that inhabit and colonise the genomes of essentially all organisms. These entities are called transposable elements. Our genome includes four million segments of DNA derived from these parasites. They comprise at least fifty percent of our DNA content. Most reproduce themselves by a copy-and-paste mechanism (DNA => RNA => DNA), and new instances are being added constantly to genomes (Figure 4).
Transposable elements reproduce in genomes haphazardly, but many are recruited to perform important functions. A study identifying some 25,000 types of transposable elements in placental mammals has described how these agents “have substantially influenced the evolutionary history of the organisms they occupy, even contributing to major evolutionary innovations.”[7] Repeatedly, sequences present in transposable elements have been coopted into the vital role of regulating gene activity. Nearly all primate-specific regulatory sequences are located within transposable elements.[8]
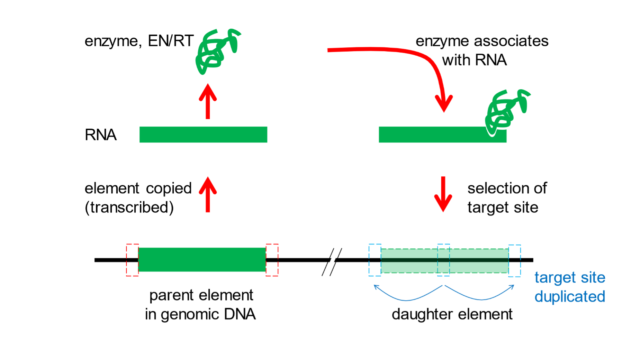
Figure 4. Mechanism by which transposons multiply in host genomes. A transposable element (green box, lower left) in the genome (horizontal line) is copied by cellular enzymes into an RNA version. The RNA provides the directions to make an enzyme that can both cut DNA (an endonuclease function, EN) and copy RNA back into DNA (a reverse transcriptase function, RT). The enzyme associates with the original RNA molecule (or sometimes, with a bystander RNA molecule), selects a target site somewhere else in the genome, cuts the DNA and inserts a DNA copy of the transposable element into the genomic DNA. During this process, the target site (dashed boxes) is duplicated on each side of the new daughter element (lower right).
(i) SVA Elements and Neurogenesis
Human brains are larger, contain more cells, and have more intercellular connections than brains of other primates. During human development, neural progenitor cells reproduce vigorously but they do not have the properties of functional neurons (nerve cells). The proliferation phase of neural progenitor cells is extended relative to those of other species, enabling the production of more brain cells. Concomitantly, the maturation of progenitor cells into nerve cells (neurogenesis) is delayed. Delayed development is called neoteny.
Transposable elements known as SVA elements, found only in great apes, are often located near genes specifying proteins with neural functions. SVA elements can bind a repressor protein called ZNF91, which silences both the SVA element and also nearby genes. ZNF91 bound to an SVA_F element that is human-specific (found only in the human genome) silences the adjacent CDK5RAP2 gene and suppresses maturation of nerve cells. When the progenitors mature into nerve cells, a distant SVA_B element is activated and generates an RNA copy, which both sequesters the ZNF91 protein, and intertwines itself into the SVA_F element. Consequently, the CDK5RAP2 gene is activated and neurons form.[9] This mechanism is shown in the upper part of Figure 5, with the insertion site of the regulatory SVA_B element being shown in the lower part.
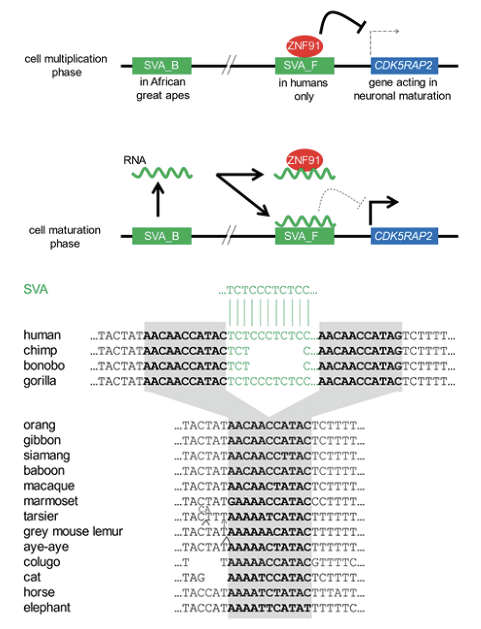
Figure 5. Involvement of an SVA transposable element in nerve cell maturation.
Above: In proliferating neural cells, a repressor molecule (ZNF91) binds to human-specific SVA elements to silence genes (such as CDK5RAP2) required for neurogenesis. At the proliferation-to-maturation decision point, an SVA element common to African great apes generates an RNA molecule that displaces ZNF91 from the human-specific SVA elements. The genes become active and neural development commences. Nadler et al. (2023). Sequences are from the UCSC Genome Browser (human); the NCBI Blast algorithm (non-human); Dfam database (SVA_B).
Below: The insertion site of the SVA_B element (green font) which generates the neurogenic RNA molecule. It entered the primate genome in an ancestor of the African great apes. The undisturbed target site is widely detectable in eutherian mammals, and its tell-tale duplications are in bold and shaded.
This example demonstrates that the African great apes (humans, chimps, bonobos, gorillas) are descendants of a common ancestor, and that SVA elements have been recruited to control genes implicated in human brain development. Human-specific SVA elements, adjacent to many genes that drive neurogenesis, may be significant contributors to human-specific brain anatomy.
(ii) Alu Elements and Adiposity
The accumulation of abdominal fat is associated with systemic (body-wide) inflammation and with chronic metabolic diseases. The deposition of abdominal fat is influenced by many genetic variants found in regulatory motifs that control genes involved in fat metabolism and adipogenesis (the development of fat cells).
Such regulatory sequences are often located in transposable elements—particularly, in Alu elements. A six-base sequence (AGGTCA; Figure 6), often present in Alu elements, recruits gene-regulating proteins that control the activity of nearby genes. An Alu element present in apes and monkeys (but not in prosimians or non-primate mammals) appears to regulate a set of nearby genes, including the NICN1 gene. Perhaps regulatory motifs in Alu elements (which are found only in primate genomes) account for the distribution of body fat that is characteristic of humans relative to other great apes.[10]
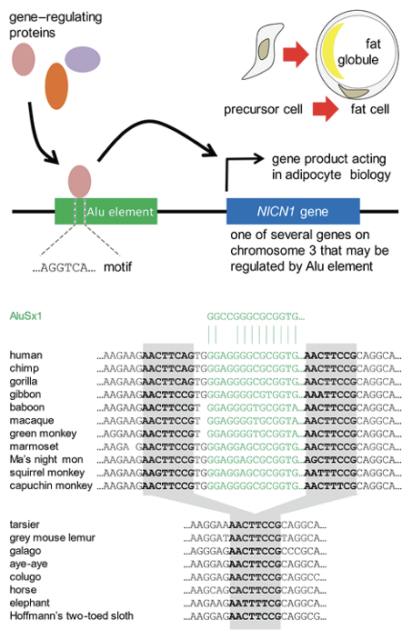
Figure 6. Involvement of an Alu transposable element in fat cell maturation
Above: An Alu element (green box) recruits proteins (ovals) that regulate adipogenesis.
Below: the insertion site in simian primates of the Alu element (green font) exerting control of the NICN1 gene (Hansen et al. 2023).
(iii) MIR Elements and Cardiovascular Disease
In heart tissue, progenitor cells may develop into either specialised heart muscle cells (cardiomyocytes) or smooth muscle cells. Maturation into heart muscle cells occurs when several gene-regulating proteins (including IRX1) are produced. The alternative pathway to smooth muscle cells is selected when an RNA molecule, containing sequences from an ancient MIRc transposable element, is generated. The MIRc sequences recruit a protein (called REST) that represses cardiogenic genes (such as IRX1). The MIRc-containing RNA achieves this by forming a triple helix with the DNA near the cardiogenic gene such that the REST protein is positioned to exert its repressive effect (Figure 7, above). Recruitment of the REST protein suppresses the IRX1 gene, as a result of which progenitor cells are directed to develop into smooth muscle cells.[11]
MIRc elements colonised genomes early in mammal evolution. The site at which this element resides (Figure 7, below) lacks target site duplications, which have been obliterated with time. The insertion site is the same for all eutherian (fully placental) mammals shown, including elephants and armadillos. The insertion event occurred in a eutherian ancestor, but the MIRc element was incorporated into the regulatory RNA only in a primate ancestor.

Figure 7. Involvement of a MIRc transposable element in smooth muscle development.
Above: Genes such as IRX1 are needed to form cardiomyocytes. An RNA molecule containing MIRc sequences recruits a repressor protein (REST) to the IRX1 gene so that smooth muscle cells are generated instead. Plaisance et al. (2023).
Below: The insertion site of the MIRc element. MIRc sequences are shown in green font. The reconstructed flanking sequence represents a sequence that might have been present in an ancestor of all eutherian mammals. However, the absence of target site duplications indicates that even this ancient sequence was markedly different from the target site into which the MIRc element was inserted in deep time.
Valuing History
It is axiomatic for Christian faith that God acts in history. The science of biological evolution seeks to elucidate the history of life—the richly complex development of Earth’s species. To complete the syllogism, evolutionary history must be a locus of God’s action, a backdrop or lead-in to God’s action in human history. It seems to follow that in a created world there should be features that are common to all histories.[12] For a start, mammalian phylogenetic trees can be depicted like those of biblical families. Figure 8 presents a partial mammalian dendrogram with the mutations described in this paper mapped on to it. It also shows a dendrogram illustrating the early history of the Hebrew people, as presented by Gordon Wenham.[13]
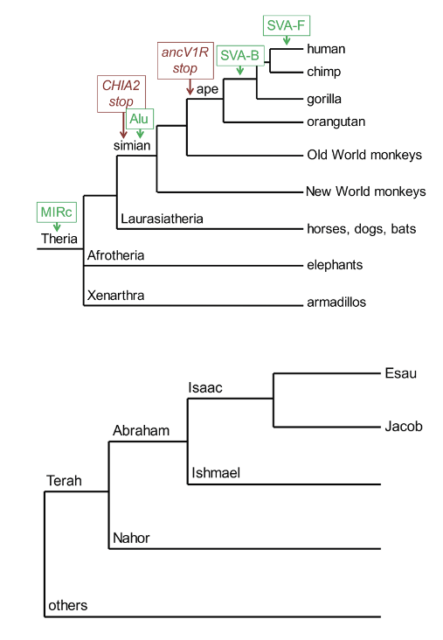
Figure 8. Partial family trees of mammals and Hebrew ancestors
Richard Dawkins’ oft-repeated assertion that evolution (biological history) enables him to be a fulfilled atheist is egregious metaphysical naivety. Christian theism is rooted in history. Dawkins’ confusion of categories can upset Christian faith only if people capitulate to the puerile presupposition that mechanistic descriptions of organic process (biological history) are a logical alternative to divine purpose and action (why there should be a biological history).
The Bible describes living organisms (including humanity and each individual), Israel and the church as divinely created.[14] But each of these created realities incorporates history as part of its being. Biological createdness encompasses a history spanning several billion years; our own created human histories, a few decades. But those histories exist by God’s will and, as aspects of the empirical world, invite investigation.
Richer Conceptions of Life
The three testaments represent stages of the one great metahistory through which the purposes of God would be achieved. They show how the phenomenon of life manifests increasing degrees of richness.
The Primal Testament, our genomic witness to biological evolution, describes the elaboration of organic life. Life is “curiously difficult to define,” but two essential features are common to all descriptions. First, a living organism is capable of reproduction, at least potentially. Second, it maintains an internal environment in which biochemical reactions occur far from thermodynamic equilibrium.[15] A living thing is a dissipative system: it maintains itself by assimilating usable (low entropy) energy and materials, and excreting them as high entropy energy (heat) and chemical waste.
The physicist Erwin Schrödinger asked the question (in 1944) “What is life?” and speculated as to the material embodiment of genetic information.[16] Schrödinger’s musings probably expedited the discovery of DNA as the carrier of genetic information (also in 1944), and the understanding of this function in chemical terms (in 1953).[17] The current paper illustrates how DNA acts as an amazingly stable but dynamically modifiable repository of life-sustaining instructions. The base sequences of DNA (Figures 2, 3, 5‒7) and the lifeforms they specify through succeeding generations describe historical process.
But a scientific understanding of life is limited to mechanism. Polkinghorne memorably said that the “world of science contains highly complicated metastable systems with the power of replicating themselves, but it does not have any people in it.”[18] Science is authoritative within its designated boundaries, but it “surveys a lunar landscape—clear, orderly, lifeless. The real gap is between talking of complex, metastable reproducing systems and talking of people.”[19]
The Old Testament provides a richer understanding of life that engages with personhood. A creature with life (hayyim) moves, and its existence is associated with light, gladness, fullness, order and activity.[20] The Hebrews used other terms also. Soul (nephesh) denoted being or self of human and non-human creatures, whether living or dead. Its “meaningful state” is living soul (nephesh hayya; Genesis 2:7). To die was to “breathe out one’s soul” (Jeremiah 15:9). Whether soul is paralleled with flesh, life, spirit, “I,” or self, the Hebrews understood that a human person was a psychosomatic unity, an animated body. The words spirit (ruach) and breath (nesama) distinguish the living from the dead and may also be used to indicate life. It follows that to die is to lose one’s spirit/breath (Psalm 104:29). Long life, a good gift from God, implied the good life (peace, prosperity, victory; Deuteronomy 30:15), but life leads inexorably to death (Sheol), at which time one’s breath returns to God, and one’s body to dust (Psalm 144:4; Genesis 3:19).
Transcending this understanding of mortal being was the conviction that Israel’s God is the source and sustainer of all life (Jeremiah 17:13). Life is lived in relatedness to God. Resurrection life was pictured in material terms—restored prosperity, peace, and fulness—although this hope was not a major theme of the Hebrew Scriptures, and its realisation belonged to the future messianic deliverance (Isaiah 26:19; Deuteronomy 12:2).
The New Testament understanding of life incorporates those features described above: organic life (Greek: bios) and soul-life (psyche), but in addition celebrates the eschatological gift of resurrection-life (res-life; one of the meanings of zoe, especially in the writings of John and Paul). Jesus’ words indicate how soul-life may be distinguished from res-life:
I have come that they might have life (zoe) and have it to the full. I am the good shepherd. The good shepherd lays down his life (psyche) for the sheep.[21]
Those who love their own life (psyche) will lose it; those who hate their own life (psyche) in this world will keep it for life (zoe) eternal.[22]
Both soul-life and res-life are the life of the self, the whole person. “The latter does not displace the former but preserves and transforms it.”[23] As in the Old Testament, life is properly the life of God, but this divine life is now manifested in Jesus[24] and concretely actualised in history.[25] In the synoptic gospels, res-life is viewed in terms of the future Kingdom of God, but in John and Paul it is also a present possession.[26]
In summary, the three testaments attest to life of different character and capacities. The Bible incorporates the scientific perspective: we are dust, chemistry, fully embedded in the natural order. As nephesh, organism, mind-body, we have continuity with the animal kingdom. But we may share in the divine life, zoe aionios, and anticipate the transformation of resurrection.[27]
Inconspicuous and Humble Origins
The assertion of human descent from ape and monkey antecedents may elicit emotive responses. But we not only share a common ancestor with apes and monkeys—biologically we are in fact apes (hominoid primates), an offshoot of the infraorder Simiiformes (monkeys or anthropoid primates). We accept that we are vertebrates and mammals, but hubris seems to make us baulk at the evidence of our more immediate affiliations with the (other) apes. We possess mutations that arose in ancestors of African great apes (Figure 5), great apes (Figure 2), anthropoid primates (Figures 3 and 6), and placental mammals that scurried around under the feet of dinosaurs (Figure 7).
We in our anthropocentric pride need to be reminded of our humble beginnings just as the ancient Israelites did. God did not love them because they were inherently important[28] or because they were particularly righteous.[29] “God loved Israel, not because they stood out as better than the rest of humanity, but precisely in spite of the fact that they exemplified (and amplified) all that is worst about humanity in general.”[30] Our value and our status arise purely from the grace of God.
The gospels describe the coming of God’s kingdom in history. As Bauckham observed, “Jesus evidently did not think that the kingdom was coming all at once. There was a process [a progressive unfolding—dare we say—an evolution]. From its small beginnings in Jesus’ own teaching and activity, the kingdom would ultimately blossom into universal reality.” God’s kingdom would start as a minute mustard seed and grow into an impressive tree. In Jesus’ own activities, “insignificant though they might seem in the grand perspective of world history, God really was sowing the seed of his coming kingdom.”[31] St Paul reminded the feuding Corinthian Christians that God “chose what the world looks down on and despises and thinks is nothing, in order to destroy what the world thinks is important.”[32] The “Christian paradox speaks of the king who reigns from the gallows.”[33]
Many people seem to be perturbed by the obscure origins and protracted meanderings of the history out of which humans arose. Such people should reflect on the equally anonymous, almost invisible histories that gave rise to God’s people in the Old Testament and to God’s newly created people in the New Testament. These histories are all of a piece. God does not love us because we are valuable; rather we are valuable because God loves us, and we rebellious apes had better get used to the fact—even as we await glorification in union with Christ.
Ambiguity
The genetic events constituting the Primal Testament inscribed in our DNA illustrate the ambiguities of our history. Our biological history is replete with amazing developments, albeit with the potential for harmful outcomes. As Tom Wright reminds us, “we are children of the present creation, which is simultaneously both glorious and shameful.”[34]
Sequences in SVA elements may help control the delayed timing of the switch in human brain cells from the proliferation phase to the maturation phase, and so allow enlarged brains. But variants in such elements are associated with mental abnormalities.[35] Sequences in Alu elements have been coopted into regulating adipocyte (fat cell) biology—but associated variants may underlie metabolic (type-2 diabetes, chronic hepatitis) and cardiovascular disease.[36] An RNA molecule containing an ancient MIRc element acts at a decision point at which progenitor cells develop into either specialised heart muscle or smooth muscle cells—but following a myocardial infarct, the MIRc-containing RNA molecule may suppress desirable myocardial cell regeneration and favour smooth muscle cell production.[37]
The Hebrew Scriptures describe how the Jewish people recognised the sea as part of God’s good creation, but at the same time, a continuing source of chaos and terror.[38] The people themselves shared in this Jekyll-and-Hyde mix of beauty and horror. Christopher Wright has said that “in the Old Testament story the people of Israel displayed a terrible ambiguity. On the one hand, they represented God’s promise to the world … But on the other hand, Israel rebelled constantly against God, and in their sin and wickedness they sank even below the level of nations that had no covenant relationship with God.” They were at the same time “a people of promise and a people in rebellion.”[39]
The third (New) Testament continues to describe the flawed characters of Jesus’ followers, but also reveals how history’s ambiguous character at last finds resolution. It is in Gethsemane and at the Cross that we may discover how God has purposed to address the sin, pain, and suffering that permeate God’s good creation. As Tom Wright says: “The multiple ambiguities of God’s actions in the world come together in the story of Jesus.[40]
Contingency
The ambiguities of history reflect their common property of contingency. The details of history do not have to turn out the way they do. We cannot predict those future events that will impact upon our lives. We are subject to happenstance. The sites at which transposable elements insert into DNA cannot be predicted and will be affected by Brownian (thermal) motion that jostles DNA templates, RNA molecules, enzymes, and carrier proteins into unpredictable configurations. When a cell divides, its daughter cells may manifest markedly different behaviour depending on how certain regulatory molecules, present in only a few copies per cell, are distributed randomly into the daughters.[41]
Histories are open to the future, reflecting the influences of spontaneous, random events. Polkinghorne said that “chance is just a shuffling mechanism for exploring potentiality” and we live in a universe of open process.[42] Osborn posits that we can describe God’s creative word in terms of promise. The structures inherent in creation manifest an open-ended “incomplete, contingent order which offers a framework for cosmic evolution.” The presence of suffering and evil suggests that the happenings in the world are not divinely micromanipulated. The promise of God represents “a commitment to the maintenance and fulfilment of its structures.”[43] God faithfully upholds the creation, and so “guarantees the freedom of the creature and the contingency of nature as a whole,” all of which underlines creation’s “radically historical” nature.[44]
Bible history also shows how the creatures act freely (so that history is contingent) even as God sovereignly achieves his ends through them. The very idea of the Covenant with Israel requires that the people were responsible for their actions. The possibility of disobedience and the derailment of God’s purposes were inherent in covenant. “The greater part of history (that of Israel, the church, the world) is in fact a violation of the will of God.”[45]
The actions of the Babylonians and Nebuchadnezzar “were wholly theirs and they bore moral responsibility” for their cruelty and violence. “God was acting through the actions of men.”[46] It is the same with the cross. Jesus was killed by men whose behaviour was motivated by an evil lust for power, and yet he died by God’s plan and foreknowledge.[47] “God acts through human actions—without turning people into puppets.”[48]
Cataclysmic Events and Extinction
We may rail against the idea of evolution because of its perceived (from our creaturely perspective) slowness, meandering character, and “wastefulness.” Every biological ancestor evinced in Figures 2, 3, 5–7 and depicted in Figure 8 is now extinct. A created order in which historical process leads to new, increasingly complex, and integrated realities is wonderfully constituted, but it “has a necessary cost not only in the blind alleys and extinctions that are the inescapable dark side of the evolutionary process, but also in the very character of the processes of a world in which evolution can take place.”[49]
The catastrophes of biological and biblical history must be faced together. The biblical testaments attest to the same pattern of haunting extinctions. Consider the meandering trajectory and apparent wastefulness of the catastrophic events that occur regularly in the Hebrew Scriptures and the New Testament. There was the destruction of Samaria (722 BC, by Assyria) and of Jerusalem (587 BC, by Nebuchadnezzar). The latter event represented the end of the kingship, the nation, the temple and its worship, and the form of the covenant which was based upon them. The cycle of cataclysms overwhelming Jerusalem continued in the centuries around the career of Jesus (167 BC, Antiochus; 63 BC, Pompey; 70 AD, Titus; 130 AD, Julius Severus). Such mass slaughter arises from the freedom exercised by sinful personal agents. We must believe that God grieves over every mutilated corpse, even as God gave solemn warning that such carnage was the inevitable result of the abuse of freedom in a world that is divinely structured according to a moral basis.
Holmes Rolston has said of evolutionary history that “the myriad creatures have been giving up their lives as a ransom for many.” The creation is cruciform.[50] So too, is God’s work in biblical (salvation) history. Transformation of the Messiah’s people by the Holy Spirit (Romans 8: 9–11) is cruciform “involving the constant life of putting to death the flesh and coming alive to the Spirit,”[51] as indeed is the experience of apostleship.[52] Overall, says Oord, God’s power to complete creation is cruciform. God will achieve God’s purposes in creation, not by overpowering and coercing the creatures, but by suffering for them.[53]
Polkinghorne acknowledges that the suffering of the creatures in both biological and biblical history can be approached by the one reality: “Much recent theological thinking has acknowledged that God’s appropriate relationship with creation includes divine suffering in compassionate solidarity with the travail of creatures. This divine passibility implies an openness to mutability,” for suffering requires the capacity to be affected by another.[54] Ultimately, people who rail against death and extinction, and the God whose rule extends over them, are railing against creaturely mortality. The story of the New Testament, dependent as it is on the resurrection of Jesus and the conferral of res-life on human creatures, has news for them. Jesus “has ended the power of death and through the gospel has revealed immortal life.”[55]
Convergence upon a Final Goal
Integral to the biblical worldview is the confidence that all histories—biological, Hebrew, or ecclesial—will culminate in a splendid climax.
The paleobiologist Simon Conway Morris has documented how diverse living forms, when faced with similar environmental challenges during phylogenetic history, frequently alight on (converge upon) similar adaptive solutions. Sharks, plesiosaurs, and dolphins have independently evolved similar streamlined shapes that confer hydrodynamic efficiency in aqueous environments. “Convergence is ubiquitous; the number of possibilities in evolution in principle is more than astronomic, but the number that actually work is an infinitesimally smaller fraction.”[56] Phylogenetic development is not a free-for-all; the structures and activities of organisms must comply with what physical law allows.
It seems that transposable elements are maintained in genomes because they provide endless variety upon which natural selection can act. Mutagenic agents are disruptive, but they work within a context of life-sustaining order. Conway Morris says, “the molecular architecture upon which mutations [the molecular dice] are imprinted—be it enzyme structure or gene networks—is rampantly convergent and greatly constrains what is possible.”[57] The phenomenon of convergence leads us to expect that “evolution perforce must follow a metaphorical map of highly restricted possibilities.”[58]
But can the tendency to progress to convergent endpoints be overwhelmed by contingent effects? Analysis of rates of genomic mutation calibrated against fossil data has shown that placental mammals diversified rapidly following the Chicxulub meteorite impact 66 million years ago. The origin of rodents and primates occurred at this time, for example.[59] However, it has been argued that extreme, random events such as bolide collisions with Earth change rates but not the overall directions of phylogenetic history. Conway Morris has said that “megadisasters such as massive flood volcanism, oceanic crises, extreme global warming, and bolide impacts undoubtedly perturb … the biosphere, but it is far less obvious that they radically redirect the course of evolution as against simply accelerating the inevitable.”[60]
The implication of ubiquitous convergence is that a creature such as Homo sapiens is a likely product of evolution. As Conway Morris says, “biology travels through history but ends up at much the same destination.”[61] So it is that “we may suppose that God can bring about determinate ends through contingent paths. Think of William James’s picture of the Grandmaster of cosmic chess, who will win the game whatever moves the creaturely opponent may make.”[62] As Polkinghorne says of cosmic history, “final fulfilment, although arrived at through the contingencies of history, is guaranteed by the steadfast love of a God ceaselessly at work within that history.”[63]
Osborn notes that as God covenants with nature, so God covenants with his people.[64] Their obedience has expected outcomes because of the moral structure of history. Their obedience to God is tantamount to a choice for life.
The biblical book of Revelation considers the painful happenstance of history in the light of the assured final victory of God. In one vision, a sealed scroll appears that “stands for the meaning and purpose of history, the great plan of God for all time.”[65] To our creaturely perspective the meaning of history is a mystery. “Who is worthy to govern history, to interpret and carry out the plan of God? … the Lamb who was slain!”[66]
Apparent chaos will indeed emerge into beauty and order. “The whole biblical narrative, at every level, is an outworking in different ways of the sovereign providence of God in the complexities of human history.”[67]
“History and theology need each other.”[68] Phylogenetic history, as much as that of Judaic and Messianic history, cries out for theological interpretation. Wright and Bird have emphasised that our contingent history will culminate in (converge upon?) a long-anticipated consummation. “Rooted in history, we are shaped by theology—not least the theological claim that God will put all things right at last” as our lives are transformed, directed to the time when “God himself, in the ultimate act of creative sovereignty, does for the whole creation what he did for Jesus when he raised him from the dead.”[69]
Final Comments
The historical sciences have revealed that we live in a universe with a fruitful history.[70] That fact should make us look beyond science as to why this should be. Polkinghorne has said that: “It is a striking fact that that initial ball of energy has become the home of saints and mathematicians. This recognition in itself might raise the question of whether there has not been more going on in cosmic history than science alone can fully express. Might there not be some purpose behind it all?”[71]
We need to reflect on the relationship between the messiness of history and the God who acts in it. This is as true of biological history as it is true of biblical history—or indeed of history at large. To Tom Wright, “The really interesting relation … is between real history in all its complexity of hypothetical reconstruction, and real faith, in all its glory as the constant exploration of, and trust in, a God who Christians believe to be … intimately and passionately involved in the historical process itself.”[72]
But what theology can we develop from impersonal and nonmoral genetic processes?
(i) If we believe we are God’s creation, then our genomic text carries the authority of that God. The Primal Testament is sacred. It is incumbent upon us to take with the utmost seriousness the historical record that we read from our DNA. During the writing of this paper, DNA sequencing studies of unprecedented scope were published. The sequencing of the genomes of 241 placental mammal species[73] and of 233 primate species[74] has led to high-level resolution of mammalian and primate phylogenetic radiations. We really do know who is related to whom. To deny that this record compels our assent, and that God is supreme over phylogenetic history, is to misrepresent the God of creation in the same way that Marcion misrepresented the God of Israel, and Arius misrepresented the triune God of Jesus’ teaching.
(ii) Our genome is a typically primate, hominoid genome. This fact is ever a corrective to hubris. Our biological rootedness explains some of our weaknesses, both physical (cardiovascular) and temperamental. Perhaps the true original sin lies in behaviours inherited from our evolutionary past, and brought into sharp relief by the advent of a moral volition and the developing awareness of God’s holy demands.[75]
(iii) Our Primal Testament reads the same way as the Old and New Testaments. The stories show inauspicious beginnings, ambiguity, contingency, cataclysm and extinction, convergence and consummation. If people had approached Darwin’s evolutionary paradigm through the spectacles of a historical framework (as described in Scripture) rather than through the spectacles of an ahistorical understanding of reality (static, made-to-order biota as described by Paley), evolution would have been accepted as a natural analogue of biblical history.[76]
Science knows nothing of teleology. It cannot in principle affirm or deny that there is a purpose to evolution. But the Primal Testament highlights the reality of convergence and supports the theological interpretation that the randomness or contingency of evolution could be expected to lead to the appearance of creatures such as we are—a primate that can hear God’s address and even feel after him.[77] Evolutionary history has in fact climaxed in a species of personal being, Homo religiosus. The Old Testament with its human history is deeply teleological. God’s distinctive promises pointed his people to the future and anticipated the unlikely climax of this history—the appearance of a saviour, a Messiah, Jesus of Nazareth.[78]
These two paradigms of realised fulfilment strengthen our confidence that the contingencies of New Testament and of church history—with their frequent failures and conflicts—will culminate in a denouement in which the people of the Messiah will witness the final establishment of God’s kingdom.
(iv) Our genetic history should inspire worship. In the incarnation, the eternal Son assumed the body, not of a demigod, but of a territorial, aggressive ape[79] in desperate need of liberation. By his total obedience to the Father, and through the scandal of the crucifixion and the shock of the resurrection, God has purposed to share the divine life and nature with his creatures.[80] By God’s grace we are more than apes, more than smart apes, more than morally aware apes. We are glorified apes, awaiting transformation.
To Polkinghorne, “our destiny beyond death is … to live an everlastingly redeemed life within the ‘time’ of the new creation. This destiny will indeed be theosis, a sharing in the life of God.”[81] To Tom Wright, the Spirit of God is again leading God’s people through the wilderness, the end result of which is theosis, being conformed to (convergent upon?) Jesus’ death and resurrection.[82]
To Osborn, the ultimate completion of the whole of created reality, is “the fulfilment of the divine promise” or “the eschatological Sabbath … [which] represents the complete participation of creation on the triune life of God.” This is the work of the Spirit of God and is cosmic theosis. Osborne sees it as involving both the nonhuman and the human creation.[83] Cosmic histories will have arrived at their single destination and “God will rule completely over all.”[84]
The author reports there are no competing interests to declare.
Received: 10/07/23 Accepted: 27/03/24 Published: 08/07/24
Appendix
A Freed Man’s Worship: The Point of It All
Quantum Vacuum
When nothing exists there’s a crowded quantum fuzz,[85]
an emptiness filled with frenetic quantum buzz;
it’s radically random with lawful compliance,
just what to expect from equations of science.
Such quantum mechanics: mere arcane abstraction,
of ceaseless invisible transient action
by quarks, leptons and bosons, weird species galore—
from such tenuous fields might we ever see more?
They blip into being, self-annihilate fast;
incessant activity—does anything last?
An ethereal gossamer-fine quantum field,
like a virtual reality—what can it yield?
Complex reality, quantum mechanics-defined,
world of a spooky, counter-intuitive kind,
must evince a creative and rational mind.
May we ask for what purpose is this brilliance destined?
Crowded quantum vacuums are lawful and are free—
might they have potential? No one but God can see.
So what, if anything, might ever be unfurled
from God’s phantasmic e’er-dependent quantum world?
Big Bang
Then from the quantum void, a shattering Big Bang,
(so some say)[86] instant world, the whole working shebang!
A cosmos emergent from the quantum vacuum,
a singularity to space-time continuum!
A fireball seeded at infinite density,
blasting through nothing to cosmic immensity.
An explosion that overcame gravity’s shackles
is evinced by pervasive CMB[87] crackles,
and hydrogen-helium ratio, three-to-one,
a proportion that serves as Big Bang’s smoking gun.
The cosmos resembles an expanding bubble;
its age is inferred from the constant of Hubble:
seventy km per sec for each megaparsec,
giving vistas both ways of this cosmic star trek.
Forces so terrible, and dimensions so vast,
a realm of pure physics, searing superhot blast;
can that fiery explosion forge anything new?
Or perchance God has plans for that dense photon brew.
Space-time’s haunting magnitude: with what is this tasked?
Without such size, duration, such questions can’t be asked.[88]
Stars
A cosmos pervaded with gas flying apart
looks so very boring—not an interesting start!
Inhomogeneities and gravity lead
to hydrogen gas falling inwards at speed.
Huge pressures ignite thermonuclear fusion,
lights piercing the blackness in hyper-profusion,
perhaps ten to the twenty-three stars, say one mole,[89]
Avogadro’s number, each one named on God’s roll.[90]
In stars, mc squared equals huge values of e—
but with ergs of such vastness can aught come to be?
A star, fuel exhausted, its lifetime being over,
may self-detonate as a bright supernova.
In its violent death-throes, it sheds ashes through space,
forming clouds of star clutter, its nebular trace.
This dust forms great shrouds that obscure cosmic features—
perhaps God will use it to make novel creatures.
Collapsed stars spawn black holes, hugely dense cosmic sinks,
with X rays emitted from their superhot brinks.
All matter is shredded in its mad inward rush—
light itself trapped in the gravitational crush.[91]
We ask what, if anything, such carnage portends:
the Creator may know, may have planned wondrous ends.
All the heavens display wondrous fireworks shows!
Is there more to this pageant? O, God alone knows.
Stars come and they go—mere cosmic futility?
Or does God’s plan entail some stellar utility?
Earth
Around a star condensing, in its accretion disk,
planets—rocks or gas globes—elliptically whisk.[92]
Many planets orbit in the Goldilocks zone,[93]
Many planets, metal-rich stardust compactions,
have chemistry giving a wealth of reactions;
there’s carbon aplenty, it’s the strangest of things,
with tetrahedrality of long chains and rings;
a CO2 blanket effects heating control,
an unseen but potent thermal greenhouse-like role.
And then there is water—a flood of surprises!
H-bonding, shows wetness, when freezing it rises.
an excellent solvent, high heat capacity:
strange curio—or from mind of sagacity?
Many planets are like this—but we know only one;
is Earth a mere odd-ball as it orbits the Sun?
It formed as hot desert, was asteroid-pelted,
had punishing windstorms,[94] volcanic rocks melted,
a speck in a cosmos of haunting dimension,
dust of no consequence—why give it a mention?
Are there prospects for Earth, roasting, sterile, so grim?
It’s all of God’s doing—we don’t know, just ask Him!
Organic Chemistry
Carbonaceous chondrites (a class of shooting star)
coat Earth with hydrocarbons (intractable tar),
and amino acids from interstellar space,
pyrogenic compounds from some cosmic cooking place.
Planet Earth a trashcan littered with amorphous junk—
is there any usefulness for aromatic gunk?
On the Earth’s deep seabed, by hot volcanic vents,
organic compounds form, endothermic events.
Can there be potential in this organic gloop?
Can anything arise from this primeval soup?
“Precursors to life!” Prebiotic chemists plead,
but they cannot imagine where such crude mixtures lead.
No one can describe how to transform random gunge
into simple animals—ctenophore or sponge.
From high temperature muck underneath the sea
can the complex components of life come to be?
O God, is there promise in this gooey thick crud?
No-one else could have schemes for organic-laced mud.
Cells
Could cells assemble by unimagined processes,
in hot, fuming darkness of undersea recesses,
with metals and sulphides, to us highly toxic,
as chemotroph feeders where all is anoxic?
Start with an RNA world! It has been opined;[95]
genetics and catalysis are here combined.
But RNA’s primary ribozyme function
is phosphodiesterase-like self-destruction.
Life is a marvel, e’en when prokaryotic;
a marvel compounded—endosymbiotic—
when free-living parents formed eukaryote cells
with specialised interdependent organelles.[96]
Acellular polymers, randomly gelling,
to self-regulation? No scheme is compelling.
The perplexities truly are far from surmounted,
the gaps in our knowledge as yet can’t be counted.[97]
Empirical science needs a hypothesis base,
a plausible route through possibility space.
Could life come by pure chance? Near impossible odds name=”_ftnref98″>[98]
without natural laws from a mind that is God’s.
If means God ordained transformed gunk to what’s living,
our only response can be gobsmacked praise-giving.
Evolution
DNA embodies genetic information.
Finely tuned stability sustains speciation:
if too high—life will forego needed adaptation;
if too low—life fails by inborn debilitation.
Variants arise from changed sequence of bases:
transposons and viruses—mutagen aces—
colonise host genomes at sites randomly placed,
gene control circuitry is facilely replaced.[99]
A crucial question: whence promoters and enhancers?[100]
Copy-paste transposons are evolution’s answers.[101]
Random mutations with strict natural selection
constrain evolution’s inherent direction;
brand new adaptations are often emergent—
but not anything goes—being widely convergent.[102]
We may wonder, amazed, but should not look askance,
at the fruitful interplay, lawfulness and chance!
This should never be controverted mystery—
authentic God creates authentic history.
Can we expect newness from constrained happenstance?
Has God charted ways through this molecular dance?
Are chance and necessity telos-embedded?
Their Author must know where phylogeny’s headed!
Bolide
Then a roadblock descends to phylogeny’s rise:
asteroid megadeath screaming in from the skies.
Iridium spike twixt K-Pg rock strata[103]
asteroid impact-incriminating data.
A layer of glass spherules from condensed molten rock,
deformed crystalline quartz, sign of high pressure shock,
two hundred km crater—Earth obliquely whacked,[104]
dinosaurs extinguished by asteroid impact,
that jolted the Earth sixty-six million years past,
nearly wiping out life by its staggering blast.
Organic forms of Earth are under cosmic fire:
are not future prospects dishearteningly dire?
Can life on Earth endure when it is bolide-zapped?
Dare we hope creation’s God has a future mapped?
Is this roasted planet, with all its sturm und drang,
the reason why the angels at creation sang?
A blast one hundred thousand billion billion joules,[105]
perhaps, just perhaps, is another of God’s tools.
Mammals
Once nuclear winter, death-dealing, had resolved,
diverse furry creatures, warm-blooded, deft, evolved.
The asteroid event was a temporary glitch,
evolution stays on track—now at a higher pitch.
The dinosaurs have gone; there is a new refrain:
it is not size that counts, but versatile brain.
Small dinosaurs called birds sing sweetly in the trees.
The platypus lays eggs, a mammal if you please,
bats like birds flap crazily, wheeling through the breeze,
cattle now called dolphins leap gladly through the seas,
bamboo-chomping pandas are vegan carnivores,
providential squirrels lay up their winter stores.
Life is so fecund, its drive autopoietic,
vigorous, exuberant! Phylogenetic
radiation leads to myriad shapes and sizes,
alpine mice to benthic whales, manifold surprises.>[106]
Chirps and caws, grunts, and barks, cacophony of praise—
do the beasts knowingly their voice of worship raise?
Is there something lacking in all creation’s throng,
creatures that freely lift to God adoring song?
Humanity
Look! A novel species of Euarchontoglires,
that makes tools for hunting, and cooks its food on fires,
useful thumb for grasping, mutant muscles in its face,[107]
furless and altricial, moves from place to place.
These brainy killer apes make war upon their kind,[108]
use tools and paint in caves, with ambivalent mind,
sometimes very tender; with kindness, love and truth,
but then sheer aggression, cruel war that knows no ruth.[109]
Are human beasts destined to pillage, kill, and rape,
that tool-using biped, most devious great ape?
The human brain does not arise by genes alone;
the social brain appears in those lovingly known.
Neural gene transcription needs personal relation,
human mind depends on love’s sure regulation.
If there’s Creating Being, of which none can conceive,
Its Word transforms life, mind, in all who dare believe.
There’s more than mere brute cunning in this hominoid:
a haunting new awareness of a God-shaped void.
Innate to the cognition of this hominin,
is instinct led by reason, wilfully, called sin.
Even now, in hedonist consumerist folly,
self-styled Homo sapiens, quintessential wally,
is trashing the planet with greed, never heeding,
crassly disregarding God’s kind, just, patient pleading.
God, what were you thinking when you brought forth this critter—
such powers, such skills, but its prospects so bitter?
When it comes to virtue it can’t do very well.
Is extinction looming, is its end soon and fell?
Is the use-by date close for woman and for man?
Or do surprises lurk in God’s mysterious plan?
Israel
From Ur, mighty city, a princeling Abram fled,
a new, inspired vision, stirring in his head:
weary of the Moon god, a new God he confessed,
who said that through his faith all people would be blessed.[110]
Through Abraham, God would address the human plight;
darkness is not forever—God will make things right.[111]
To the Hebrew folk by cruel slavery broken,
Moses, leading them, God’s Covenant had spoken:
“I will be your God; my people you will be.”[112]
Obey my holy Law and then you will be free.
A Covenant of grace—see God’s will embedded!
YHWH and his bride in joy were truly wedded.[114]
David sang, the Lord’s my shepherd, I will have no needs,
He comforts, waters, cares; and he most surely feeds.
But soon the people spurned God’s loving wise commands;
Solomon the Stupid ignored God’s just demands:
eschew Egyptian horses, great wealth, pagan brides,[115]
choose YHWH, not idols—you simply must take sides!
When true worship falters, catastrophe runs rife,
nation sinks in turmoil, injustice grows, and strife;
Jeremiah thunders, the people block their ears,
place their hope in armies, the surest way to tears.
Jerusalem is razed, the Covenant is shredded,
to exilic torment the people now are headed.
Think upon the future—could New Covenant arise
from Israel’s creator, God of sea and skies?
Could minds and hearts express a law that is God’s own?
From the smallest to the greatest, could Godself be known?[116]
Could thoughts of redemption and forgiveness be pulsing
through traumatised hearts when the world is convulsing?
Has the Hebrew project come to end in acrid smoke?
Were all of YHWH’s promises just a bitter joke?
Oppressed by cruel Babylon, Hellenisers, Rome—
has YHWH turned away forever from his home?
Exile—six hundred years—with racked anticipation,[117]
the faithful wait yearning for God’s emancipation.
Dare they hope for freedom and that YHWH has planned
to bring life to dry bones and a desolate land?
Was God’s pledge to humanity just a sick dream,
Israel, channel of blessing? A parched and dried-up stream!
The great Hebrew story: does it lie dead in the dust,
their history so dark, that we can’t sanely trust
there is hope of a sequel and that God has it sussed?
Can beauty come from ashes[118]>—please act God, you must!
Jesus
Then in the most radiant, mellifluous morning
comes Jesus announcing the joyous new dawning;
God’s kingdom is imminent,[119] hope rises anew,
is this God’s great moment, please what will God do?
He welcomed the outcasts, poor, lepers, lame, and blind,
gentiles, women, children, not one was left behind.
He soon provoked fury: “the riff-raff don’t belong
to YHWH’s holy kingdom—rabbi, you’ve got it wrong!”
Jesus and his friends came at Passover, to dine
on icons of himself, familiar bread and wine;
to release sun’s trapped energy, grain must be crushed,
to free up solar power stores, grapes must be mushed,
to liberate God’s life-force, remove human guilt,
Jesus’ body must be broken, his life-blood spilt.
Holy loving teacher, now bloodied, bruised, and torn,
on two rough-hewn wood beams, reviled in deepest scorn,
bearing Israel’s anguish, abandoned and forlorn—
is darkness back forever, with no hope of morn?
The One who’s the good shepherd, light, truth, living bread,
is killed by a crazed mob, a joke crown on his head.
Many people thought he would set Israel free;
this hope seems extinguished by the gore on the tree—
from a callous spear-thrust, blood and water released,[120]
his heart has stopped beating, truly he is deceased.
Jesus’ autolysed corpse moulders in a dark tomb;
surely nothing’s forthcoming from cold silent gloom.
Can even God see past the rank horror of death?
Can even God’s Spirit instil vital new breath?
To cultured despisers, a dead crim on a cross
is laughingstock only, a pathetic dead loss,
devoid of all value, lowest scum of the earth—
God, don’t you like winners, surely this farce lacks worth?
Must nature and humans, as if under a curse,
face only extinction in a heat-dead universe?
New Creation
Then on Easter Sunday a brand new world appears,
death and sin are vanquished, the end of human fears!
Now Jesus has been raised, his body is transformed;
every proud and hateful stronghold truly has been stormed.
Evil, death and sorrow do not have the final say;
God will make all things new![121] Rejoice in the new day!
Maria, first to hear the Lord’s voice, precious sound,
he called her Miriam, her true self had been found.[122]
Cleopas and his mate, whose grieving minds were churning,
heard Jesus on the road, with joy their hearts are burning.[123]
Thomas scorned his colleagues’ tale with incredulity;
seeing Jesus was the first to see his deity.[124]
Unprecedented sign! What’s the indication?
Jesus has the Father’s utter vindication.
Jesus resurrected? Are nature’s laws deranged?
The world is recreated, ontology is changed!
God is always faithful: this world’s laws aren’t repealed;
but in resurrection, the new world’s laws, revealed.
The law of the Spirit supersedes the law of death,[125]
eternal life is here, infused by God’s own breath.
The strife of cosmic history makes brilliant sense at last,
creation’s winding journey, its travail, pain, now past;
cross and resurrection unleashed life-giving force
from God into creation to chart a whole new course.
Church
The church bursts into life—no languid evolution—
the worth of all affirmed, the Christian revolution,
proclaiming to all folk, peace, justice, charity,
sexes, races, classes, one in perfect parity.
God’s mercy reaches all, there is not one exception,
we’re equal in God’s sight—true human rights inception.[126]
The church must serve in love that humankind be blest,
and that God’s healing power will truly be confessed.
And yet the church so fails God’s grace to manifest,
therefore, ecclesia semper reformanda est.[127]
United in love, we proclaim Whom we follow;
with rancour, division, that rings very hollow.
The church is so plagued by abundance of “isms”—
its witness is damaged by fragmenting schisms.
Feebly we model Your indiscriminate grace,
Your surpassing mercy to each child of each race;
O Lord, has the plan for your world been aborted?
Your righteous demands have been widely distorted.
We say we’re disciples—but, recalcitrant breed,
cannot extirpate the rampant selfishness weed;
Your people sink helpless in a morass of greed,
so small our compassion for your poor in great need;
we so blindly pursue the consumerist creed—
is there hope in our time? Lord please speak, help us heed.
Science
In tribal myth—spirits, gods, trees, sky are blended,
and should not be disturbed—the gods might be offended.
For Hebrew tribes only, creation’s transcended,
de-deified; God’s world may be comprehended.[128]
The gods of the Greeks are conniving, capricious;
but Israel’s God, sovereign, truthful, judicious.
Consistent and lawful, the world God created:
a gift of the Gospel—science is gestated![129]
The Stoic world cycles through endless reversions,
so lassitude undermines research exertions.
But the God who redeems acts through history’s flows,
the cause-and-effect linear time that science knows.[130]
God’s nature is goodness, and God’s world must be good;
our glad confidence in the world’s preciousness would
enhance nature’s beauty before wondering eyes,
to aid and abet growth of the science enterprise.
God made nature in freedom, the world is contingent,
so researchers explore, humbly and with most stringent
empiricality and hypothesis testing
to reveal nature’s laws, the prized fruit of their questing.
“The glory of God and the good of humanity,”
underlie science; which may sink to inanity
by the pursuit of prestige and of monetary gain;
science is God’s gift; is divine largesse in vain?
Lord you are the Truth, and when you are rejected,
integrity needed for science is neglected.[131]
When deaf to your Logos, existential thought rules,
we create our own “truth”—will we wink out like fools?[132]
Conspiracy theories pollute even God’s church;
Where’s the Spirit of Truth? Are we back in the lurch?
Addicted to falsehood, reckless ideation,
dare we hope “God will not abandon creation”?[133]
Eschaton
Quantum void, hot Big Bang, stellar fires—God ordained,
desert Earth, complex goo, simple cells—God maintained,
life, beasts and then people, Israel, church—God sustained,
in them God’s plan was gestated, good purpose contained.
Now God’s kingdom’s concealed, it seems merely latent;
then wondrous surprise, God’s just rule will be patent.
Now beauty is haunting, and we cannot hold it;
then beauty is lasting, with joy we’ll enfold it.
Whilst first sight of mountain, forest, coastline enthrals,
with subsequent seeing, the high ecstasy palls;
but Father-Son glory, creation transcending,[134]
we’ll witness enraptured, in awe never-ending.
Lord, your ways with people, show uniformity—
not of despotic might—of cruciformity.
New Creation’s cost to you, profound enormity;
till then, help us follow you in full conformity.
You make things of wonder from the crudest of dross,
the supreme paradigm, that most hideous cross;
to us, cogent symbol of love fully expressed,
of your mercy supreme, grace by which we are blessed.
As players in your saga of suffering to glory,[135]
we’ll see the denouement of your cosmic story.
Your plan for the ages, from mute matter begun,
is to fill New Creation with folks like your Son![136]
The point of the journey, this painful biosis,
imbued with your life by the Son’s great kenosis,[137]
to arrive at a creaturely apotheosis:
a dull primate enwrapped into loving theosis!
The Spirit’s vitality, end of all thirsting,
the spring of the water of life, gushing, bursting
forth, filling us, as the pure source of our being,
with your truth, fully known, with acuity seeing.
In full obedience, Jesus went to the cross,
to offer all people zoe aionios;
for groaning creation, he’s Omega, Telos,
the true goal and purpose, the final Eschatos.[138]
The most wondrous secrets inhere in the Trinity,
we’ll learn, rejoice, worship through time’s great infinity.
To wonder at those eternal scars on the Lamb
To rejoice in the love of the triune I AM.
[1] John Polkinghorne, Science and the Trinity (New Haven: Yale University Press, 2004), 38‒39.
[2] The word testament is used in the same way in each of the three divinely written histories to indicate that they are ordained, sustained, and consummated by the will of God (cf. Hebrews 9:15‒18).
[3] Francis S. Collins, The Language of God (New York: Free Press, 2006).
[4] Cristina Sisu, “GENCODE Pseudogenes,” Methods in Molecular Biology 2324 (2021): 67–82, DOI: 10.1007/978-1-0716-1503-4_5; or see pseudogene.org.
[5] Hikoyu Suzuki et al., “A Single Pheromone Receptor Gene Conserved Across 400 Million Years of Vertebrate Evolution,” Molecular Biology and Evolution 35 (2018): 2928–2939, DOI: 10.1093/molbev/msy186.
[6] Christopher A. Emerling et al., “Chitinase Genes (CHIAs) Provide Genomic Footprints of a Post-Cretaceous Dietary Radiation in Placental Mammals,” Science Advances 4 (2018): eaar6478, DOI: 10.1126/sciadv.aar6478.
[7] Austin B. Osmanski et al., “Insights Into Mammalian TE Diversity through the Curation of 248 Genome Assemblies,” Science 380 (2023): eabn1430, DOI: 10.1126/science.abn1430.
[8] Gregory Andrews et al., “Mammalian Evolution of Human Cis-Regulatory Elements and Transcription Factor Binding Sites,” Science 380 (2023): eabn7930, DOI: 10.1126/science.abn7930. These authors showed that for 367 gene-regulating DNA binding proteins, the genome had 15.6 million binding sites, of which 3.8 million were found only in primates, and 3.3 million overlapped transposable elements.
[9] Monica J. S. Nadler et al., “Hominoid SVA-LncRNA AK057321 Targets Human-Specific SVA Retrotransposons in SCN8A and CDK5RAP2 to Initiate Neuronal Maturation,” Communications Biology 6 (2023): 647, DOI: 10.1038/s42003-023-04683-8.
[10] Grace T. Hansen et al., “Genetics of Sexually Dimorphic Adipose Distribution in Humans,” Nature Genetics 55 (2023): 461–470, DOI: 10.1038/s41588-023-01306-0.
[11] Isabelle Plaisance et al., “A Transposable Element into the Human Long Noncoding RNA CARMEN is a Switch for Cardiac Precursor Cell Specification,” Cardiovascular Research 119 (2023): 1361–1376, DOI: 10.1093/cvr/cvac191.
[12] As has been argued: Graeme Finlay, “Evolution as Created History,” Science and Christian Belief 20 (2008): 67‒89; Evolution and Eschatology (Eugene: Wipf and Stock, 2021), 142‒158; “Evolution as History: Phylogenetics of Genomes and Manuscripts,” Christian Perspectives on Science and Technology, New Series 1 (2022): 150‒174, DOI: 10.58913/JJHH2131.
[13] Gordon Wenham, Exploring the Old Testament Volume. 1: The Pentateuch (London: SPCK, 2003), 46.
[14] For example, Genesis 1:21; Psalm 104:30; Genesis 1:27; Psalm 139:13; Isaiah 43:1, 15; Ephesians 2:15.
[15] Christopher Southgate et al., God, Humanity and the Cosmos (Edinburgh: T&T Clarke, 1999), 144.
[16] See Rob Phillips, “Schrödinger’s What is Life? at 75,” Cell Systems 12 (2021): 465–476, DOI: 10.1016/j.cels.2021.05.013.
[17] As updated for the Watson-Crick-Wilkins-Franklin structure of DNA. See Matthew Cobb and Nathaniel Comfort, “What Watson and Crick Really Took from Franklin,” Nature 616 (2023): 657–660.
[18] John Polkinghorne, The Way the World Is (London: SPCK, 1983), 16.
[19] John Polkinghorne, One World (London: SPCK, 1986), 60.
[20] J. D. Douglas, ed., The Illustrated Bible Dictionary (Leicester: IVP, 1980), 901–902.
[21] John 10:10–11. See in Bernard Palmer, ed., Medicine and the Bible (Exeter: Paternoster, 1986), 133.
[22] John 12:25.
[23] Douglas, Illustrated Bible Dictionary, 903.
[24] Most explicitly in the Gospel of John and the letters, as follows: John 1:4; 5:26; 14:6; 1 John 5:20.
[25] 1 Corinthians 15; Colossians 3:4.
[26] Douglas, Illustrated Bible Dictionary, 904. See Mark 10:17; Matthew 25:46 cf John 5:24; 1 John 3:14; Ephesians 2:4.
[27] Donald M. MacKay, The Clockwork Image (London: IVP, 1974), 70–71.
[28] Deuteronomy 7:7–8.
[29] Deuteronomy 9:4–6.
[30] Christopher J. H. Wright, The God I Don’t Understand (Grand Rapids: Zondervan, 2008), 113–115.
[31] Richard Bauckham, Jesus: A Very Short Introduction (Oxford: Oxford University Press, 2011), 37.
[32] 1 Corinthians 1:28, GNT.
[33] Polkinghorne, The Way the World Is, 77.
[34] N. T. Wright, Surprised by Scripture (London: SPCK, 2014), 201‒202.
[35] Nadler et al., “Hominoid SVA-Lnc RNA,” 647.
[36] Hansen et al., “Genetics of Sexually Dimorphic Adipose Distribution,” 461‒470.
[37] Plaisance et al., “Transposable Element into the Human,” 1361‒1376.
[38] Wright, Surprised by Scripture, 116.
[39] Wright, The God I Don’t Understand, 145–146 (italics original).
[40] Wright, Surprised by Scripture, 120.
[41] As Phillips points out, “partitioning errors in transcription factors are a demonstrable part of the reason for noisy gene expression” (“Schrödinger’s What Is Life? at 75,” 470).
[42] Polkinghorne, Science and the Trinity, 78–79.
[43] Lawrence Osborn, Guardians of Creation (Leicester: IVP, 1993), 114.
[44] Osborn, Guardians, 115.
[45] Adrio Konig, Here Am I: A Believer’s Reflection on God (Grand Rapids: Eerdmans, 1982), 130.
[46] Wright, The God I Don’t Understand, 147.
[47] Acts 2:23.
[48] Wright, The God I Don’t Understand, 147.
[49] Polkinghorne, Science and the Trinity, 72.
[50] Holmes Rolston III, Genes, Genesis and God (Cambridge: Cambridge University Press, 1999), 306–307.
[51] N. T. Wright, Paul and the Faithfulness of God (London: SPCK, 2013), 1023.
[52] 2 Corinthians 4—6.
[53] Thomas J. Oord, The Uncontrolling Love of God (Downers Grove, IL: IVP, 2015), 155.
[54] Polkinghorne, Science and the Trinity, 109.
[55] 2 Timothy 1:10.
[56] Simon Conway Morris, The Runes of Evolution (Conshohocken: Templeton, 2014), 21.
[57] Conway Morris, Runes, 22.
[58] Conway Morris, Runes, 24.
[59] Nicole M. Foley et al., “A Genomic Timescale for Placental Mammal Evolution,” Science 380 (2023): eabl8189, DOI: 10.1126/science.abl8189.
[60] Conway Morris, Runes, 22.
[61] Conway Morris, Runes, 6.
[62] Polkinghorne, Science and the Trinity, 108.
[63] John Polkinghorne, Science and Christian Belief (London: SPCK, 1994), 66.
[64] Osborn, Guardians, 90–91, 94–95.
[65] Revelation 5 and succeeding chapters that describe the opening of the seals.
[66] Wright, The God I Don’t Understand, 64.
[67] Wright, The God I Don’t Understand, 97–98.
[68] N. T. Wright and Michael F. Bird, The New Testament in its World (London: SPCK, 2019), 81.
[69] Wright and Bird, New Testament in its World, 83.
[70] Polkinghorne, Science and the Trinity, 65f.
[71] Polkinghorne, Science and the Trinity, 66.
[72] N. T. Wright, Jesus and the Victory of God (London: SPCK, 2016), 8–9.
[73] Foley et al., “Genomic Timescale,” esp. Figure 1.
[74] Lukas F. K. Kuderna et al., “A Global Catalog of Whole-Genome Diversity from 233 Primate Species,” Science 380 (2023): 906–913, DOI: 10.1126/science.abn7829; Yong Shao et al., “Phylogenomic Analyses Provide Insights into Primate Evolution,” Science 380 (2023): 913–924, DOI: 10.1126/science.abn6919.
[75] Polkinghorne, Science and Christian Belief, 15.
[76] As indicated by Peter Harrison in Karl W. Giberson, ed., Abraham’s Dice (Oxford: Oxford University Press, 2016), 278–279.
[77] Acts 17:27.
[78] 1 Peter 1:10–12, 20.
[79] José María Gómez et al., “The Phylogenetic Roots of Human Lethal Violence,” Nature 538 (2016): 233–237, DOI: 10.1038/nature19758.
[80] For example, John 3:14–16; 6:53–58; 11:25–26; 2 Peter 1:4.
[81] Polkinghorne, Science and the Trinity, 115–116.
[82] N. T. Wright, Paul and the Faithfulness of God (London: SPCK, 2013), 1023.
[83] Osborn, Guardians, 127.
[84] 1 Corinthians 15:28; see also Romans 8:18–25; Ephesians 1:10; Col 1:20.
[85] It has been said that Hawking described the cosmos “like a kind of fuzzy spacetime egg”; biblically, the Creator is to be understood “as the sustainer of the self-contained spacetime egg and as the ordainer of its quantum laws” (Polkinghorne, Science and Christian Belief, 73).
[86] In quantum field theory, there “are no particles present, no permanent excitations, but there is a continual chaos of transient blips … On this view we might be living in a grotesquely swollen quantum fluctuation,” an idea that Polkinghorne describes as highly speculative. See John Polkinghorne, Science and Creation (London: SPCK, 1988), 58–60.
[87] CMB, cosmic microwave background, discovered as static on a Bell Labs radio antenna (Arno Penzias and Robert Wilson, 1965).
[88] It takes a multibillion year cosmic history to make humans, says Polkinghorne: “Without all those trillions and trillions of stars, we should not be here to be dismayed by them” (One World, 57). And Davison adds, “A connection between size and age [of the universe] and the possibility of the formation both of elements and of stars with their solar systems, is well established.” Andrew Davison, Astrobiology and Christian Doctrine (Cambridge: Cambridge University Press, 2023), 93.
[89] In chemistry, one mole is 6.023 x 1023 atoms or molecules, and is of a similar order of magnitude to the estimated number of stars in the universe.
[90] Isaiah 40:26; Psalm 147:4.
[91] As told entertainingly by Heino Falcke, Light in the Darkness (London: Wildfire, 2021), 92–95.
[92] The James Webb Space Telescope is providing unprecedented insights into planetary formation. See the news items by Alexandra Witze, “Life in the Cosmos: JWST Hints at Lower Number of Habitable Planets,” Nature (19 June 2023), https://www.nature.com/articles/d41586-023-01983-1; “How Worlds are Born: JWST Reveals Exotic Chemistry of Planetary Nurseries,” Nature (20 September 2023), https://www.nature.com/articles/d41586-023-02922-w.
[93] Of the order of 1019 according to Andrew Davison, Cambridge Professor of Theology and the Natural Sciences, https://biologos.org/podcast-episodes/andrew-davison-cosmic-incarnation (accessed 30 November 2023).
[94] Early Earth rotated faster than it does now (having slowed by interaction with the Moon), generating hurricane-force winds. See Simon Conway Morris, Life’s Solution (Cambridge: Cambridge University Press, 2003), 90.
[95] Francisco Prosdocimi and Savio Torres de Farias, “Origin of Life: Drawing the Big Picture,” Progress in Biophysics and Molecular Biology 180–181 (2023): 28–36.
[96] It has been proposed that endosymbiosis underlies many phases in life’s history. See Predrag Slijepcevic, “Serial Endosymbiosis Theory: From Biology to Astronomy and Back to the Origin of Life,” Biosystems 202 (2021): 104353.
[97] Charles Kocher and Ken A. Dill, “Origins of Life: First Came Evolutionary Dynamics,” QRB Discovery 4 (2023): e4, 1–9, DOI: 10.1017/qrd.2023.2.
[98] A thought adapted from Jim Brooks, Origins of Life (Tring: Lion, 1985), 87 and 103; this delightful book excited my interest nearly forty years ago.
[99] For example, retrotransposons have influenced brain development. See Raquel Garza et al., “L1 Retrotransposons Drive Human Neuronal Transcriptional Complexity and Functional Diversification,” Science Advances 9 (2023), eadh9543; Alexander Frohlich et al., “CRISPR Deletion of a SINE-VNTR-Alu (SVA_67) Retrotransposon Demonstrates its Ability to Differentially Modulate Gene Expression at the MAPT Locus,” Frontiers in Neurology 14 (2023), 1273036, DOI:10.3389/fneur.2023.1273036.
[100] Promoters are DNA motifs adjacent to genes upon which the enzymatic machinery responsible for gene expression is assembled. Enhancers are motifs widely scattered through the genomes, that control the cell type, time, and intensity of gene expression.
[101] Liang Liang, Changchang Cao, Lei Ji et al. “Complementary Alu Sequences Mediate Enhancer–Promoter Selectivity,” Nature 619 (2023): 868–875, DOI: 10.1038/s41586-023-06323-x.
[102] For a summary of the significance of evolutionary convergence, see Simon Conway Morris, “The Paradoxes of Evolutionary Convergence,” Faraday Paper 26 (2022), https://www.faraday.cam.ac.uk/wp-content/uploads/2022/09/Faraday-Paper-26-Conway-Morris.pdf.
[103] K-Pg, Cretaceous-Paleocene (previously, K-T, Cretaceous-Tertiary) boundary.
[104] Gareth S. Collins et al., “A Steeply-Inclined Trajectory for the Chicxulub Impact,” Nature Communications 11 (2020), article no. 1480, DOI: 10.1038/s41467-020-15269-x.
[105] Joanna V. Morgan et al., “The Chicxulub Impact and its Environmental Consequences,” Nature Reviews Earth & Environment 3 (2022): 338–354.
[106] See the news report by Elizabeth Pennisi, “Mountain Mice Break Altitude Record for Mammalian Life,” Science 382 (2023): 360.
[107] MYH16 pseudogene, specific to humans. See George H. Perry et al., “Insights into Hominin Phenotypic and Dietary Evolution from Ancient DNA Sequence Data,” Journal of Human Evolution 79 (2015): 55–63, DOI: 10.1016/j.jhevol.2014.10.018.
x[108] Polkinghorne, The Way the World Is, 19.
[109] Hunter-gatherer settlements in Siberia show evidence of violent conflict (palisade protection, destruction by fire, projectile heads). See Henny Piezonka et al., “The World’s Oldest-Known Promontory Fort: Amnya and the Acceleration of Hunter-Gatherer Diversity in Siberia 8000 years ago,” Antiquity 97 (2023): 1381–1401, DOI: 10.15184/aqy.2023.164.
[110] Genesis 12:3; 18:18; 22:18.
[111] “God had chosen Israel … to be the means of rescuing humans and the whole creation” (Wright and Bird, The New Testament in Its World, 511; also 259).
[112] Adrio Konig, The Eclipse of Christ in Eschatology (Blackwood: New Creation, 1999), 54, 82, 95; 240; and see Leviticus 26:12; Hosea 2:23; Jeremiah 7:23; 11:4; 24:7; 30:22; 31:1; 31:33; 32:38; Ezekiel 11:20; 14:11; 37:23; 37:27; Zechariah 8:8; 13:9; 2 Corinthians 6:16; Romans 9:25–6; Hebrews 8:10; Revelation 21:3.
[113] Exodus 34:6; Psalm 103:8; 145:8–9
[114] Hosea 2:14–23.
[115] Deuteronomy 17:16–17; cf. 1 Kings 10:14—11:13; 2 Chronicles 2: 14–17; 8:11; 9:13–28.
[116] Jeremiah 31:31–34; for the sequel, Hebrews 8:7–13.
[117] The Jews considered their exile to continue, even after returning to their own land, as they faced continued oppression. See N. T. Wright, The New Testament and People of God (Minneapolis: Fortress, 1992), 268–272.
[118] Isaiah 61:3.
[119] Mark 1:15; Matthew 4:17, 23; Luke 4:16–21, 47; John 3:3, 5.
[120] John 19:34; “blood,” sedimented erythrocytes, “water,” cell-free plasma, indicating decomposition of blood.
[121] Revelation 21:5.
[122] John 20:11–18; Mary Magdalene was usually called by her name in Greek, Maria; but when she met Jesus resurrected, he addressed her by her birth name, Miriam. See Wright, Surprised by Scripture, 216.
[123] Luke 24:13–35, esp. 14, 32.
[124] John 20:24–29.
[125] Romans 8:2.
[126] David Bentley Hart, Atheist Delusions (New Haven: Yale University Press, 2009), 166–182.
[127] “The church must always be reformed.”
[128] Harold Turner, The Roots of Science (Auckland: DeepSight Trust, 1998), 54–78.
[129] Mark Worthing, Unlikely Allies: Monotheism and the Rise of Science (Eugene: Wipf & Stock, 2019); Peter Harrison, “Religion and the Rise of Science,” Faraday Paper 21 (2021), https://www.faraday.cam.ac.uk/wp-content/uploads/2021/03/Faraday-Paper-21-Harrison-web.pdf.
[130] Stanley L. Jaki, Science and Creation (Edinburgh: Scottish Academic Press, 1986), 102–137.
[131] Scientific integrity is being eroded by corrupt practices. Perhaps 400,000 papers published over the last twenty years (up to 2% of the whole) are fraudulent. See Richard Van Noorden, “How Big is Science’s Fake-Paper Problem?” Nature 623 (2023): 466–467.
[132] For a comment on the dangers of disinformation to human survival, see Volker Turk, “Protect the ‘Right to Science’ for People and the Planet,” Nature 623 (2023): 9.
[133] A hope-filled assertion given by Professor Sir John Houghton at a public lecture on climate change I attended, Auckland.
[134] John 17:4–5, 24.
[135] John 12:27; Luke 24:26; Acts 3:13, 18; Romans 8:18; Hebrews 2: 9, 10; 1 Peter 1:11; 4:13; 5:1, 10.
[136] A big-picture summary learned from my Father Matthew.
[137] Philippians 2:7.
[138] Revelation 1:17; 2:8; 22:13.